ORIGINAL RESEARCH ARTICLE
Infiltration level of macrophages and its impact on fat graft retention in mouse free fat grafting model
Chengzhi Donga  , Jian Zhangb and Lijuan Penga
, Jian Zhangb and Lijuan Penga
aDepartment of Plastic Surgery, Tongde Hospital of Zhejiang Province, Hangzhou, China; bThe Second Clinical Medical College, Zhejiang Chinese Medical University, Hangzhou, China
ABSTRACT
Background: Autologous fat grafting faces the clinical challenge of unstable absorption rates, and the regulatory mechanisms of macrophages need to be elucidated.
Objectives: It aimed to investigate the infiltration levels and phenotypic dynamics of macrophages and explore their impact on graft retention rate (RR).
Methods: A total of 180 C57BL/6J mice were randomly divided into experimental (AG, fat grafting), sham (BG, no grafting), and intervention (CG, macrophage depletion followed by grafting) groups. Graft RR was assessed, and macrophage status was analyzed by flow cytometry and immunofluorescence postoperatively. Levels of inflammatory, pro-fibrotic, and angiogenic factors were also detected. Results: Macrophage infiltration in the AG peaked at week 2 (21.3 ± 1.5%) and then shifted from M1 to M2 phenotype. The graft RR in the AG was visibly higher as against the CG (p < 0.05). M2 macrophages were strongly positively correlated with RR (based on the aggregated data of all time points, r = 0.821, p < 0.0001), while pro-inflammatory factors suggested negative correlation. The AG had visibly elevated levels of angiogenic factors, which were strongly positively correlated with RR (p < 0.05).
Conclusion: Macrophages (especially the M2 subtype) play a key role in fat graft survival by modulating inflammatory responses and promoting angiogenesis. Targeting macrophage polarization may be a novel strategy to improve fat grafting outcomes.
KEYWORDS: fat grafting; macrophage polarization; graft retention rate; angiogenesis; inflammatory response
Citation: Journal of Plastic Surgery and Hand Surgery 2025; 61: 95–103. DOI: https://doi.org/10.2340/jphs.v61.45744.
Copyright: © 2026 The Author(s). Published by MJS Publishing on behalf of Acta Chirurgica Scandinavica. This is an Open Access article distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), allowing third parties to copy and redistribute the material in any medium or format and to remix, transform, and build upon the material, with the condition of proper attribution to the original work.
Received: 30 October 2025; Accepted: 27 February 2026; Published: 5 May 2026.
CONTACT: Lijuan Peng 1730156947@qq.com Department of Plastic Surgery, Tongde Hospital of Zhejiang Province, No. 234, Gucui Road, Hangzhou City, Zhejiang Province, China
Competing interests and funding: The author declares that there are no financial or non-financial conflicts of interest.
This research received grant from Zhejiang Medical and Health Science and Technology Program Project 2024KY870.
Aim
This study aims to systematically evaluate the infiltration level of macrophages and their subtype dynamics in transplanted tissues and clarify the impact of macrophages on tissue retention rate at different time points after fat grafting by establishing the mouse free fat grafting model. It also further explores the potential mechanisms that may be mediated by macrophages (such as inflammatory response, angiogenesis, and tissue remodeling), providing a theoretical basis and intervention targets for improving the stability and long-term effects of fat grafting.
Core tips
This study confirmed through the mouse model that the dynamic infiltration of macrophages (especially M2-type polarization) after fat grafting significantly enhances graft survival through a dual-mechanism approach: on the one hand, by secreting factors such as vascular endothelial growth factor to promote angiogenesis and improve graft blood supply; and on the other hand, by regulating anti-inflammatory factors such as interleukin-10 to reduce local inflammatory responses. This finding provides a new intervention idea and therapeutic target for the clinical improvement of fat grafting success rate by targeting macrophage polarization (such as promoting M2-type transformation).
Introduction
With the development of plastic surgery and regenerative medicine, autologous fat grafting is widely adopted for soft tissue defect repair and cosmetic reshaping due to its good biocompatibility, abundant source, and natural tissue-building ability. However, high absorption rates and unstable volume retention after fat grafting seriously affect its clinical efficacy and expected outcomes. Sustained retention rate (RR) of transplanted fat varies visibly, ranging from 20 to 80% [1, 2]. Therefore, in-depth analysis of the core mechanisms affecting the survival and stability of free fat grafts is urgently needed in the clinic to optimize grafting techniques and improve long-term efficacy.
Previous studies have shown that the survival of free fat grafts is highly dependent on the re-vascularization of the graft and the regulation of the host immune microenvironment [3, 4]. Research on the mechanisms of inflammatory responses related to fat grafting has gradually become a hot topic. Relevant studies have found that macrophages widely infiltrate the graft in the early stages of transplantation, participate in the regulation of inflammatory responses and tissue remodeling, and are a key immune cell population connecting innate immunity, angiogenesis [5], and tissue repair [6]. Macrophages have visible plasticity and can be polarized into pro-inflammatory M1 and anti-inflammatory/reparative M2 subtypes under the influence of the local microenvironment [7]. Among them, M1 macrophages can exacerbate local tissue damage and fat necrosis by secreting inflammatory factors [8], while M2 macrophages promote fat re-vascularization and repair [9]. The dynamic changes and polarization direction of the M1/M2 macrophage ratio in the early microenvironment of fat grafting may directly affect the survival quality and RR of the graft [10].
Although previous studies have preliminarily revealed the infiltration and distribution characteristics of macrophages in fat-grafted tissues, their role in the early stages of transplantation is still a matter of debate. On the one hand, moderate infiltration of M1 macrophages can induce the clearance of necrotic tissue and activate subsequent repair mechanisms. On the other hand, excessive M1 response or delayed M2 response may lead to aggravated tissue necrosis and fibrosis formation [11]. In addition, the impact of macrophages on angiogenesis has also attracted much attention. They can regulate the formation of new blood vessels and thus affect the nutrient supply and metabolic waste clearance efficiency of the graft [12]. Therefore, systematically exploring the infiltration levels and polarization states of macrophages in the early stages of fat grafting from a dynamic and functional perspective will help to reveal their specific roles in the regulation of fat RR. Based on the background of this study and the unresolved debates, the following hypotheses were proposed: (1) The dynamic polarization of infiltrating macrophages from a pro-inflammatory (M1) to a pro-repair (M2) phenotype positively correlated with fat graft RR; (2) Macrophages, particularly the M2 subtype, enhanced graft survival by modulating the local inflammatory response and promoting angiogenesis in the early post-transplantation period.
To validate these hypotheses, a murine free fat graft model was established in this study, and methods such as immunohistochemistry, flow cytometry, and quantitative analysis of fat RR were used to systematically evaluate the infiltration density, subtype composition of macrophages, and their correlation with angiogenesis, tissue necrosis, and RR at different time points after fat grafting. The goal is to reveal the key role of macrophages in the immune microenvironment of fat grafting and provide a basis for potential immune-regulatory strategies to improve the clinical success rate of fat grafting.
Materials and methods
Experimental animals
A total of 180 male C57BL/6J mice, aged 6–8 weeks and weighing 20–25 g, were purchased from the Shanghai Laboratory Animal Center. The mice were housed under standard conditions at the Laboratory Animal Center, with a 12-hour light-dark cycle, 22 ± 2℃, and relative humidity kept at 55 ± 10%. During the experimental period, the animals had free access to standard laboratory animal feed and were provided with sufficient drinking water. Protocols obtained the approval by the Animal Ethics Committee, and strictly adhered to the related guidelines to ensure animal welfare and the scientific nature of the experiments.
Grouping method
A simple randomization method was adopted. After numbering 120 mice, a random sequence was generated using a computer-based random number generator (Excel random function). According to the sequence, the mice were divided into the following groups.
Experimental group (AG, n = 60): autologous free grafting of inguinal fat pads.
Sham group (BG, n = 60): only skin incision and no fat grafting.
Intervention group (CG, n = 60): the donor fat was treated with clodronate liposomes to deplete macrophages before transplantation.
The grouping process was completed by an independent experimenter to avoid subjective interference from the operator. After grouping, each group was randomly assigned to different subgroups in proportion according to postoperative time points (1, 2, 4, 8 weeks) (n = 15 per subgroup) to ensure balanced baseline characteristics among subgroups. The grouping information was kept confidential until data analysis, and both the experimental operations and outcome assessments were performed in a blinded manner. Following group assignment, baseline measurements and comparisons of age and weight were performed for all mice to verify inter-group homogeneity. The ages of mice in the AG, BG, and CG groups were (7.1 ± 0.6) weeks, (7.0 ± 0.7) weeks, and (7.2 ± 0.5) weeks, respectively, while their weights were (22.3 ± 1.4) g, (22.5 ± 1.6) g, and (22.1 ± 1.5) g, respectively. One-way analysis of variance (ANOVA) revealed no statistically significant differences among the three groups in either age (p = 0.852) or weight (p = 0.736), indicating effective randomization, balanced baseline characteristics across groups, and a reliable experimental design.
Fat tissue collection and processing methods
Fat collection: One day before surgery, the mice were anesthetized with isoflurane (induction 3%, maintenance 2%, RWD Life Science (RWD)-3000 small animal anesthesia system, RWD Life Science, China). After anesthesia, the mice were placed on a sterile operating table, where the inguinal area was shaved and disinfected successively with 75% ethanol (Sinopharm Chemical Reagent Co., Ltd.) and 2% iodophor (Runying Medical, Jiangsu, China). Then, sterile ophthalmic scissors and toothless forceps (Fine Science Tools, Germany) were adopted to open the skin and fascia, and the bilateral inguinal fat pads were carefully dissected. The fat pads were immediately placed in pre-cooled sterile phosphate-buffered saline (PBS) (Gibco, 10010-023, USA) after complete dissection and temporarily stored in a 1.5 mL sterile centrifuge tube (Axygen, USA) (4°C). The entire collection process was completed within 5 minutes to maximize the quality of the fat tissue.
Macrophage depletion treatment: The fat tissue from the CG required macrophage removal treatment. The freshly collected inguinal fat pads were placed in a sterile solution containing clodronate liposomes (purchased from Liposoma BV, Netherlands, final density 2 mg/mL) (water bath, 37℃, 1 h). The centrifuge tubes were gently shaken during incubation to ensure full exposure of the fat tissue to the drug solution. After incubation, the fat tissue was washed three times with sterile PBS (Gibco, Cat#10010-023) (each time for 5 minutes) to remove residual drugs and avoid their impact on the animals. The treated fat tissue was immediately adopted for subsequent transplantation.
Fat grafting procedure: All mice were anesthetized, disinfected, and a 1-cm incision was made locally in the skin with a sterile surgical blade (Feather No.15, Feather Safety Razor Co., Japan), followed by gentle dissection of the subcutaneous tissue to create a potential cavity. The pre-treated fat tissue (about 0.3 mL) was slowly injected into the cavity using a 1-mL disposable sterile syringe (BD, USA, 27G needle). The incision was then continuously sutured with 5-0 absorbable sutures (Ethicon Vicryl Rapide, USA) and coated with an antibacterial ointment (erythromycin ointment, CR Sanjiu Medical) to prevent infection. The control group only underwent skin incision and cavity creation at the corresponding site without fat implantation, followed by similar suture of the incision. All postoperative mice were placed on a warming pad to recover and returned to the breeding cage, with continuous observation for 72 h for any postoperative infection, abnormal activity, or wound dehiscence.
Tissue collection and preservation
At 1, 2, 4, and 8 weeks postoperatively, mice were euthanized by cervical dislocation, and the skin was quickly cut open to completely remove the fat grafts. According to the requirements of subsequent tests, tissue samples were placed under different preservation conditions: fresh tissues for flow cytometry were placed in pre-cooled PBS; tissue blocks for immunofluorescence and frozen sectioning (-80℃ after Optimal Cutting Temperature (OCT) compound embedding); tissue blocks for Enzyme-Linked Immunosorbent Assay (ELISA) and molecular detection (-80℃); tissues for histological analysis were subjected to fixation in 4% paraformaldehyde and subsequently preserved by paraffin embedding. At week 8, the complete grafts were taken out, and the graft weight and micro-Computed Tomography (CT) scanning were immediately performed after drying the surface moisture.
Flow cytometry analysis
To quantitatively analyze the infiltration level of macrophages and the distribution of M1/M2 subtypes in the fat graft area, single-cell suspensions were prepared from about 50 mg of fat tissue from the graft area. The minced tissues were placed in a mixed digestion solution of collagenase type I (1 mg/mL, Sigma, USA) and DNase I (20 µg/mL, Roche, Switzerland), digested in a water bath (37℃, 45 minutes), filtered through a 70-µm mesh (Corning, USA), and then centrifuged and subjected to resuspension in PBS. After cell counting, the concentration was adjusted to 1 × 10⁶ cells/100 µL, incubation with Fc receptor-blocking solution (BioLegend, USA) (10 minutes) and co-incubation with a mixture of antibodies CD45-APC (eBioscience, USA), F4/80-PE, CD86-FITC, and CD206-PerCP-Cy5.5 (all purchased from BioLegend, USA) (30 minutes, 4℃, in the dark). After washing with PBS, dead cells were excluded with 7-AAD. Data were collected using a BD LSRFortessa flow cytometer (Biosciences, USA), with no less than 20,000 CD45+ events recorded for each sample. The gating strategy in FlowJo v10.8 software (v10.8, BD Biosciences, USA) was CD45+ → F4/80+ → CD86+/CD206+, adopted to analyze the total amount of macrophages and the proportion of subtypes.
Immunofluorescence staining
To further observe the spatial distribution of macrophages in the fat graft tissue, about 100 mg of fat tissue was taken for immunofluorescence staining analysis. The preserved tissues were sectioned into 8-µm slices with the relative device (CM1950, Leica Microsystems, Germany) and mounted on poly-l-lysine-coated slides (Citoglas, China). Following air-drying (25℃), PBS was adopted for rinsing, 1% BSA + 0.3% Triton X-100 (Sigma-Aldrich, USA) was adopted for blocking, and the primary antibody F4/80 (ab6640, Abcam, UK, 1:200) was applied (overnight, 4℃). The next day, following rinsing, the secondary antibody labeled with Alexa Fluor 594 (Invitrogen, USA, 1:500) was applied (25℃, 1 h), and the nuclei were counterstained with DAPI (Thermo Fisher Scientific, USA, 1 µg/mL). After staining, the sections were mounted with Fluoromount-G (SouthernBiotech, USA), and observation was carried out adopting the relative device (Axio Imager M2, Zeiss, Germany). Five high-power fields (400×) were randomly selected from each section for counting F4/80+-positive cells to assess the infiltration level of macrophages in the fat tissue.
Graft RR assessment
Fresh fat grafts were taken, and the graft weight was measured with an electronic balance (Sartorius, Germany, precision 0.1 mg) after gently drying the surface moisture with filter paper. The fat RR was calculated according to the equation: RR = (Weight taken out / Initial graft weight) × 100%. Micro-CT scanning (Skyscan 1176, Bruker, Belgium) was then performed. Before scanning, iopromide contrast agent (Ultravist 370, Bayer, Germany) was injected through the tail vein to enhance the visualization of the fat. The Micro-CT scan data in Digital Imaging and Communications in Medicine (DICOM) format was imported into the VG Studio MAX 3.0 software (Volume Graphics, Germany) for volumetric analysis. Firstly, grayscale calibration was performed on the images to standardize brightness and contrast. Subsequently, a semi-automatic threshold segmentation method was applied, setting a Hounsfield Unit (HU) range of −200 to −50 to distinguish adipose tissue from the surrounding soft tissues. Based on the segmentation results, a three-dimensional model was generated. The ‘Region Growing’ tool was then used to refine the edges and remove noise. Finally, the software automatically calculated the volume (unit: mm³) of the segmented region, and the data were exported for the calculation of graft volume RR. A detailed schematic of the process is presented in Figure 1.
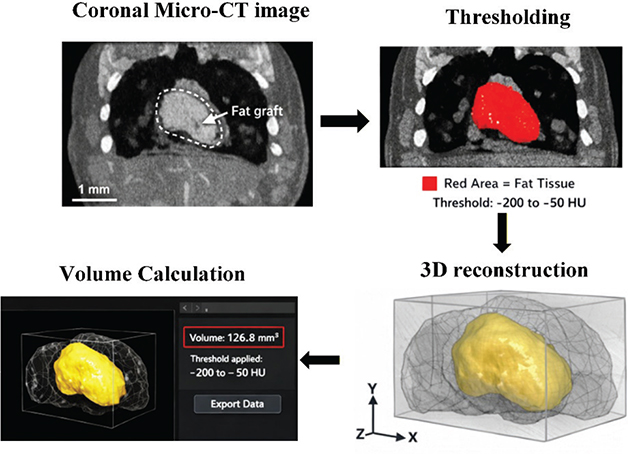
Figure 1. Flowchart of volume analysis using VG studio MAX 3.0.
Detection of inflammatory and angiogenic factors
The tissue samples preserved in Section 2.4 were homogenized in PBS at a 1:5 ratio, and the supernatant was taken for detection after centrifugation. Tumor necrosis factor-α (TNF-α), interleukin (IL)-6, and IL-1β were detected using a mouse ultra-sensitive multi-factor detection kit (BioLegend, USA) according to the kit requirements. Transforming growth factor-β (TGF-β) was detected using an acid-activated ELISA kit (Research and Development (R&D) Systems, USA), which was treated with 1 M HCl for 10 minutes before sampling and then neutralized with an alkaline buffer. Type I collagen was detected using a pro-collagen peptide detection kit (Cloud-Clone Corp., China) after pepsin (Sigma-Aldrich, USA) pre-treatment. Vascular endothelial growth factor (VEGF) and fibroblast growth factor 2 (FGF-2) were detected using conventional single-factor ELISA kits (Abcam, UK). All samples were measured at 450 nm wavelength, and calculation was carried out in combination with the standard curve, with concentrations represented as pg/mg protein. The detection of each factor was performed according to the specific requirements of the corresponding ELISA kit.
Statistical methods
All data were statistically analyzed adopting GraphPad Prism 9.0 (GraphPad Software, USA) or SPSS 26.0 (IBM, USA). Continuous variables were represented as mean ± SD. Two-way repeated-measures ANOVA was adopted for multi-group and multi-time point data, with Bonferroni method for multiple contrast; independent-samples t-test or Mann–Whitney U test was adopted for single-measurement contrast, depending on normality and homogeneity of variance. One-way ANOVA was adopted for contrast among multiple groups, and Tukey’s Honestly Significant Difference (HSD) test was applied for pairwise contrast. Correlation analysis was performed adopting Pearson or Spearman method. The distinction was statistically considerable with p < 0.05. To ensure statistical power, sample size was determined based on pilot experiments and literature data through power analysis, with no less than 6–8 mice per group.
Results
Macrophage infiltration levels and subtype dynamics
(1) Flow cytometry analysis
The proportion of macrophages (CD45+F4/80+) in the graft area of the AG visibly increased to 15.8 ± 1.2% at 1 week postoperatively, which was elevated as against the BG (4.3 ± 0.7%, p < 0.01). It peaked at 2 weeks (21.3 ± 1.5%) and then gradually decreased to 18.7 ± 1.3% at 4 weeks and 14.9 ± 1.1% at 8 weeks. Subtype analysis suggested that the proportion of M1 macrophages (CD86+) was 42.5 ± 3.6% at 1 week postoperatively and decreased to 35.8 ± 2.9% at 2 weeks. In contrast, the proportion of M2 macrophages (CD206+) continuously increased from 38.2 ± 3.1% to 56.4 ± 4.2% at 8 weeks. The macrophage infiltration level in the CG was visibly lower as against the AG at all time points (p < 0.01), and the subtype distribution was mainly M1, lacking the dynamic upregulation trend of M2 (Figure 2).
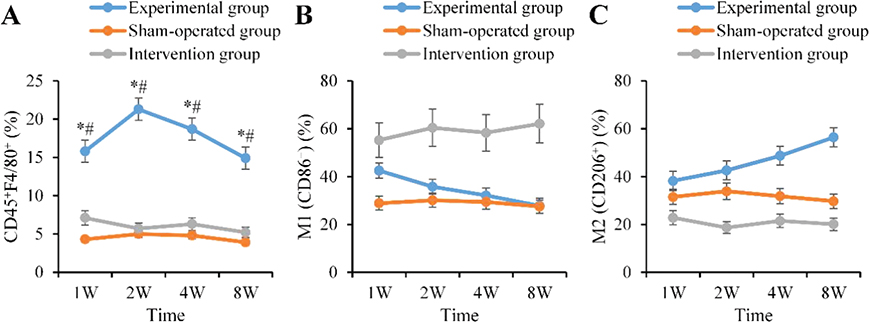
Figure 2. Proportion of macrophages and M1/M2 subtypes in the fat graft area (%). (A: Total macrophages B: M1 type C: M2 type ‘*’: AG vs. BG, ‘#’: AG vs. CG, p < 0.05).
(2) Immunofluorescence spatial distribution and quantitative results
Immunofluorescence quantitative results suggested that the number of F4/80+ macrophages in the AG was 35.6 ± 3.2 per high-power field at 1 week postoperatively, which was visibly elevated as against the BG (8.9 ± 1.5) and the CG (12.4 ± 2.0) (both p < 0.05), and further increased to 48.2 ± 4.1 at 2 weeks. Although there was a slight decrease at 4 weeks (42.7 ± 3.6) and 8 weeks (36.4 ± 3.0), it remained higher as against the BG (7.8 ± 1.3 and 6.5 ± 1.2) and the CG (11.2 ± 2.1 and 9.8 ± 1.5) (p < 0.05). No statistical distinction was found between the BG and the CG (p > 0.05). Spatially, the AG group exhibited extensive distribution of bright red signals (F4/80+ cells), particularly forming aggregations around blood vessels and within adipose lobular septa. In contrast, the red signals in the BG group were sparse and diffusely scattered. While the graft structure remained visible in the CG group, its red signal intensity was markedly lower than that in the AG group (Figure 3).
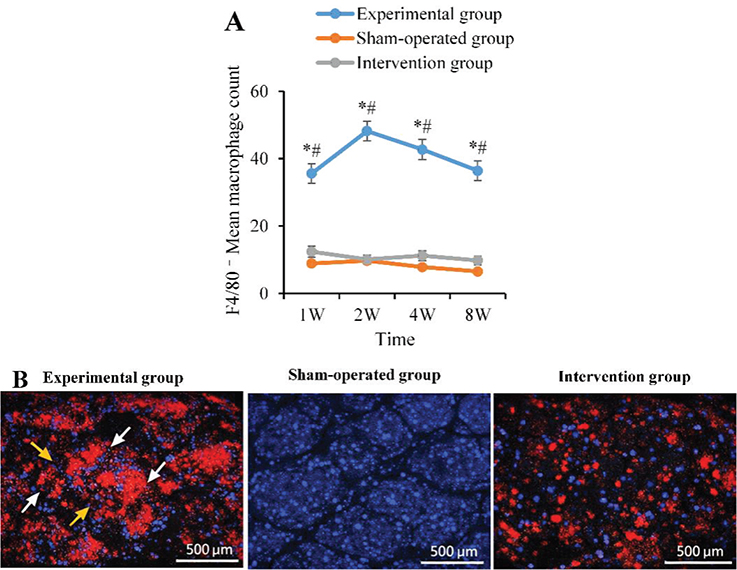
Figure 3. Average count of F4/80+ macrophages in the graft (per high-power field). (A: Quantitative results of F4/80+ macrophages; B: Immunofluorescence staining images at 2 weeks post-operation (100×), with white arrows indicating areas of macrophage aggregation; yellow arrows indicating perivascular infiltration. ‘*’: AG vs BG, ‘#’: AG vs. CG, p < 0.05).
Fat graft RR and micro-CT volume analysis
The fat RR gradually decreased over time after surgery, but the AG was always visibly better than the CG. The weight RR of the AG at 1, 2, 4, and 8 weeks postoperatively were 91.3 ± 3.2%, 85.9 ± 4.6%, 76.5 ± 6.0%, and 68.7 ± 5.8%, respectively; the volume RR was 93.4 ± 2.9%, 87.2 ± 3.5%, 78.0 ± 5.2%, and 65.1 ± 6.3%, respectively. In contrast, the weight RR of the CG decreased to 84.7 ± 4.1%, 71.2 ± 5.7%, 52.3 ± 6.4%, and 38.9 ± 7.2%, and the volume RR was also visibly lower (86.1 ± 3.7%, 70.4 ± 5.0%, 49.5 ± 5.9%, and 36.2 ± 6.7%, respectively). The distinctions between the two groups were statistically meaningful from 2 to 8 weeks postoperatively (p < 0.05). Furthermore, representative macroscopic photographs of the explanted fat grafts demonstrated that, at 8 weeks postoperatively, the grafts in the AG group exhibited a larger volume and intact morphology, whereas those in the CG group showed significant atrophy with an irregular surface (Figure 4).
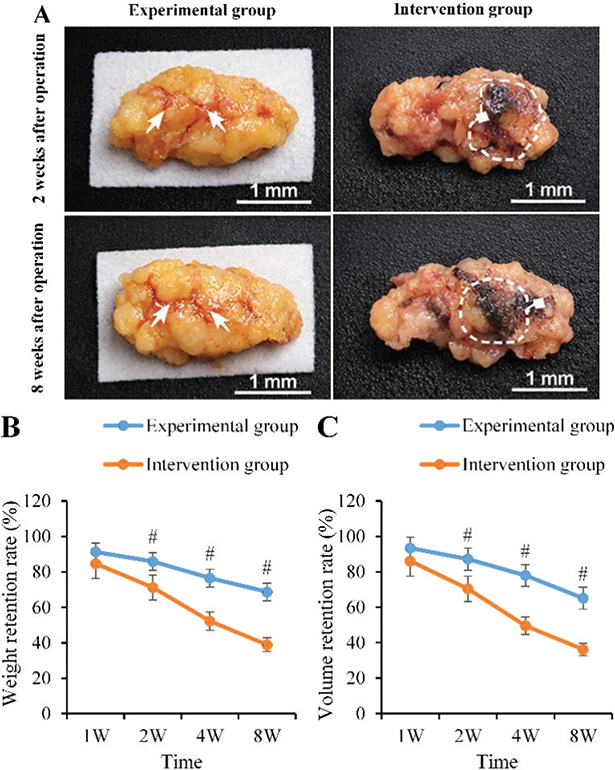
Figure 4. Comparison of fat graft RR. (A: Macroscopic photographs of explanted samples from each group at 2 and 8 weeks post-operation, wherein the white arrows indicate the graft boundary and areas of angiogenesis; areas marked by dashed boxes represent regions of graft atrophy, fibrosis, or necrosis.); B: Weight RR; C: Volume RR; ‘#’: AG vs CG, p < 0.05). RR: retention rate.
Analysis of inflammatory factor changes
The AG peaked at 1 week postoperatively (TNF-α: 120.5 ± 15.3 pg/mg; IL-1β: 85.4 ± 9.8 pg/mg; IL-6: 96.7 ± 11.2 pg/mg), which were visibly higher as against the BG (p < 0.05). They began to decline at 2 weeks (78.6 ± 8.9 pg/mg; 52.1 ± 6.3 pg/mg; 60.3 ± 7.1 pg/mg) and returned to baseline levels by 8 weeks. In contrast, those in the CG abnormally increased at 1 week (210.8 ± 20.1 pg/mg; 156.2 ± 14.3 pg/mg; 158.9 ± 16.5 pg/mg) and remained highly expressed at 2 weeks (192.4 ± 18.5 pg/mg; 140.8 ± 13.7 pg/mg; 130.5 ± 12.8 pg/mg), and were visibly higher as against the AG until 8 weeks (p < 0.05) (Figure 5).
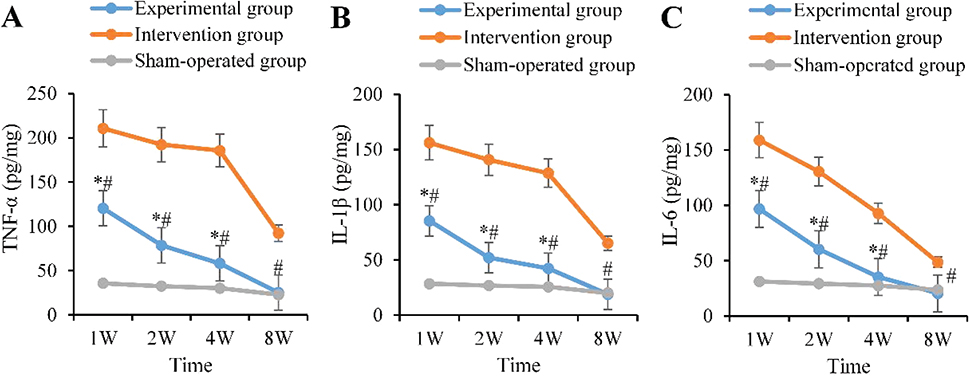
Figure 5. Dynamic changes of pro-inflammatory factors. (A: TNF-α B: IL-1β C: IL-6; ‘*’: AG vs. BG, ‘#’: AG vs. CG, p < 0.05). TNF-α: tumor necrosis factor-α; IL-1β: interleukin-1β; IL-6: interleukin-6.
Analysis of pro-fibrotic factor changes
Pro-fibrotic factors in the AG progressively increased. At 1 week postoperatively, they were at low levels (TGF-β: 32.6 ± 4.1 pg/mg; Collagen I: 18.3 ± 2.4 pg/mg), markedly rising from 1 to 2 weeks (48.7 ± 5.8 pg/mg; 30.5 ± 3.9 pg/mg), peaking at 4 weeks (65.3 ± 7.2 pg/mg; 42.1 ± 5.1 pg/mg), and partially declining by 8 weeks. In the CG, the levels at 2 weeks were markedly lower as against the AG (35.2 ± 4.3 pg/mg vs. 48.7 ± 5.8 pg/mg; 21.7 ± 2.8 pg/mg vs. 30.5 ± 3.9 pg/mg), and the 4-week peak was still lower (41.7 ± 5.3 pg/mg; 26.4 ± 3.6 pg/mg) (p < 0.05), only approaching the levels in the AG by 8 weeks (Figure 6).
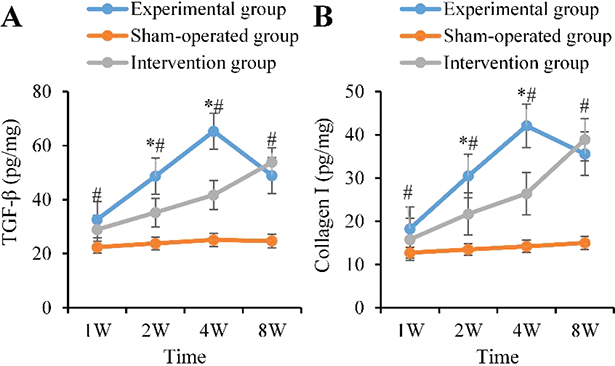
Figure 6. Dynamic changes of pro-fibrotic factors. (A: TGF-β B: Collagen I; ‘*’: AG vs BG, ‘#’: AG vs CG, p < 0.05). TGF-β: transforming growth factor-β.
Analysis of angiogenic factor changes
In the AG, the levels of angiogenic factors VEGF and FGF-2 markedly increased from the 2nd week, peaked at the 4th week (VEGF: 38.7 ± 4.5 pg/mg, FGF-2: 29.8 ± 3.6 pg/mg), and remained at medium-high levels at the 8th week. The levels at 2, 4, and 8 weeks were markedly elevated as against the BG (p < 0.05). In the CG, VEGF and FGF-2 were always markedly lower as against the AG. The peaks at 4 weeks were even less than 50% of those in the AG (19.2 ± 2.3 pg/mg vs. 38.7 ± 4.5 pg/mg; 14.9 ± 1.9 pg/mg vs. 29.8 ± 3.6 pg/mg, p < 0.01), and they returned to near pre-operative degrees by 8 weeks (Figure 7).
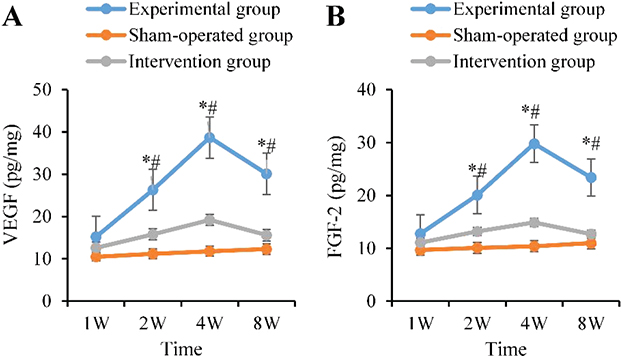
Figure 7. Dynamic changes of angiogenic factors. (A: VEGF B: FGF-2; ‘*’: AG vs. BG, ‘#’: AG vs. CG, p < 0.05). VEGF: vascular endothelial growth factor; FGF-2: fibroblast growth factor 2.
Correlation analysis between fat RR and microenvironmental factors
To clarify the relationship between fat RR and local microenvironmental factors, the data from all postoperative time points (1, 2, 4, and 8 weeks) were integrated using Spearman’s correlation analysis. The results suggested that fat RR was moderately positively correlated with the total amount of macrophages (r = 0.647, p = 0.0014), with the proportion of M2 type having the strongest correlation (r = 0.821, p < 0.0001), while the proportion of M1 type was negatively correlated (r = -0.583, p = 0.0038). Inflammatory factors were all markedly negatively correlated with RR (r ranged from -0.733 to -0.669, p < 0.001). TGF-β and Collagen I were moderately positively correlated with RR (r = 0.516, 0.482, p < 0.05). VEGF and FGF-2 were highly positively correlated with RR (r = 0.836, 0.769, p ≤ 0.0001) (Table 1).
Discussion and conclusions
This study established a mouse model of free fat grafting and systematically analyzed the dynamic infiltration and subtype polarization of macrophages post-transplantation. It was found that macrophage infiltration was significantly elevated in the early stage and gradually shifted toward M2-type polarization. Depletion of macrophages not only reduced their overall quantity and blocked M2 polarization but also led to a decrease in fat RR, while simultaneously exacerbating pro-inflammatory responses and inhibiting pro-fibrotic and angiogenic processes. Correlation analysis indicated that the fat RR was positively correlated with M2-type macrophages, pro-fibrotic factors, and angiogenic factors, and negatively correlated with M1-type macrophages and pro-inflammatory factors.
Our findings revealed that macrophages within the fat grafts of the experimental group were rapidly recruited, reaching a peak at 1–2 weeks post-operation, a trend consistent with previous studies. For instance, Lin et al. [13] observed a significant increase in macrophage numbers by the second week after fat grafting, which aligns with the graft's early requirement to cope with ischemia-reperfusion injury and necrotic tissue clearance [14]. Further analysis demonstrated that macrophages in the experimental group gradually shifted from an early M1-dominant state toward M2 polarization, with the M2 proportion exceeding 50% by the 8th week. This finding was further supported by research from Chen et al. [15]. This polarization process conforms to the ideal repair mechanism following tissue injury: early M1 macrophages clear necrotic debris and trigger inflammation, while the subsequent emergence of M2 macrophages helps to suppress inflammation, promote angiogenesis, and facilitate tissue remodeling [7, 16]. In contrast, the intervention group, where macrophages were depleted, disrupted this dynamic polarization process, leading to insufficient M2 conversion, which in turn resulted in sustained high inflammation, impaired angiogenesis, and decreased tissue retention.
The results from the intervention group (macrophage depletion group) were highly instructive. As anticipated, macrophage depletion significantly reduced the RR of the fat grafts. Notably, macrophage depletion did not alleviate the early post-operative (1–2 weeks) inflammatory response; instead, it led to abnormally elevated levels of pro-inflammatory factors (TNF-α, IL-1β, IL-6) with delayed resolution. This phenomenon suggests a complex dual role of macrophages in the transplant microenvironment: (1) macrophages themselves, particularly the M1 subtype, are a major source of inflammatory factors; (2) however, their timely polarization to the M2 phenotype is crucial for clearing necrotic cells and terminating inflammation [17, 18]. Our findings support the predominant role of the latter mechanism, indicating that macrophage depletion impedes necrotic tissue clearance, leading to the continuous release of ‘danger signals’ that activate an uncontrolled inflammatory cascade, thereby exacerbating tissue damage [19]. This mechanism explains the aggravated inflammatory response and the decreased graft RR observed in the intervention group.
Furthermore, the expression of pro-fibrotic factors (TGF-β, Collagen I) and angiogenic factors (VEGF, FGF-2) in the intervention group was significantly delayed and reduced, particularly during the critical 2–4 week time window. Macrophages, especially the M2 subtype, are important sources of these reparative factors [20, 21]. TGF-β and moderate collagen deposition (Collagen I) form the basis for tissue scaffold reconstruction and fibrous capsule formation, contributing to graft stabilization [22, 23]. VEGF and FGF-2 are key drivers of graft revascularization [24, 25]. The absence of macrophages directly weakened the initiation and intensity of these repair and regeneration processes, resulting in insufficient graft revascularization, inadequate nutrient supply, accumulation of metabolic waste, and ultimately accelerating adipocyte apoptosis, necrosis, and resorption. The strong positive correlations observed in the correlation analysis between fat RR and the proportion of M2 macrophages, VEGF, FGF-2, and TGF-β further support the synergistic role of these factors in maintaining graft survival.
In summary, this study demonstrated that targeted regulation of macrophage polarization holds promise for improving the repair efficiency and long-term survival rate of fat grafts by optimizing the immune microenvironment. However, several limitations remain: Firstly, the conclusions are derived from a mouse model, and differences in immune response and fat metabolism between mice and humans necessitate caution when directly extrapolating findings to the clinical setting [26]. Secondly, to control variables and reduce individual differences, only male mice were used. Given that sex hormones are known to influence immune responses and fat metabolism, the generalizability of our findings to female individuals requires further validation. Future studies should include female animals to comprehensively assess the impact of sex differences. Thirdly, the specific molecular mechanisms by which macrophages regulate angiogenesis and fibrosis, such as the roles of key signaling pathways (e.g. PI3K/Akt, STAT), remain unclear and warrant further investigation using methods such as conditional gene knockout or cell co-culture experiments. Fourthly, while the clodronate liposome depletion method used effectively reduced macrophages, it may non-specifically affect other myeloid cells (e.g. monocytes, some dendritic cells). Furthermore, this method cannot distinguish between tissue-resident macrophages and those derived from recruited monocytes, and functionally important differences may exist between these subsets. Fifthly, the observation period of this study was limited to 8 weeks post-operation, primarily reflecting mid-term changes in graft survival and remodeling. An assessment of long-term dynamics (e.g. beyond 6 months) regarding macrophage behavior, fat tissue stability, fibrosis outcomes, and final volume RR is lacking.
In conclusion, macrophages play a pivotal role in reshaping the microenvironment of free fat grafts. Moderate infiltration and timely polarization toward the M2 phenotype help control inflammation, promote angiogenesis and tissue repair, thereby ensuring graft survival. Conversely, macrophage depletion disrupts immune balance and reduces survival rates. These results suggest that optimizing fat grafting should focus on regulating macrophage polarization and function, rather than simply inhibiting their infiltration.
Authors’ contributions
Conceptual design: Chengzhi Dong, Lijuan Peng; Research methods: Chengzhi Dong, Jian Zhang; Data collection and analysis: Chengzhi Dong, Jian Zhang; the first draft of the article was written by: Chengzhi Dong; Article revision and review: All authors; Funding support: Lijuan Peng; Resource provision: Lijuan Peng; Research supervision: Lijuan Peng.
Acknowledgments
This study was supported by the Zhejiang Medical and Health Science and Technology Program (Grant No. 2024KY870). The authors would like to thank the Laboratory Animal Center of Zhejiang Chinese Medical University for their technical assistance and support.
Ethics declarations & trial registry information
This study was approved by the Animal Ethics Committee of [Institution Name] (Approval No. [number]). All animal procedures were performed in accordance with the relevant guidelines and regulations. This study did not involve human subjects; therefore, informed consent was not applicable.This study was a preclinical animal experiment and was not registered in a clinical trial registry.
Data availability statement
The data generated and analyzed in this study can be obtained from the corresponding author upon reasonable request.
References
[1] Ørholt M, Weltz TK, Hemmingsen MN, et al. Long-term volume retention of breast augmentation with fat grafting depends on weight changes: a 3-year prospective magnetic resonance imaging study. Plast Reconstr Surg. 2025;155(6):947–954. https://doi.org/10.1097/PRS.0000000000011841
[2] Thomas B, Warszawski J, Falkner F, et al. Fat grafts show higher hypoxia, angiogenesis, adipocyte proliferation, and macrophage infiltration than flaps in a pilot mouse study. Plast Reconstr Surg. 2023;152(1):96e–109e. https://doi.org/10.1097/PRS.0000000000010146
[3] Yeğin EE, Yeğin ME, Kosova B, et al. Analysis of fat graft survival and platelet-rich plasma effects: the transcriptomic differences. Cureus. 2023;15(1):e34380. https://doi.org/10.7759/cureus.34380
[4] Chen G, Long J, Zhang Y, et al. Fascia-derived stem cells enhance fat graft retention by promoting vascularization through the HMOX1-HIF-1α pathway. Stem Cell Res Ther. 2025;16(1):92. https://doi.org/10.1186/s13287-025-04204-w
[5] Bi X, Li B, Zou J, et al. Fascia promotes adipose tissue regeneration by improving early macrophage infiltration after fat grafting in a mouse model. Plast Reconstr Surg. 2023;152(3):446e–457e. https://doi.org/10.1097/PRS.0000000000010259
[6] Wang Y, Guo X, Zeng H, et al. M2 macrophage polarization and tissue remodeling in autologous fat grafting for diabetic skin defects. J Craniofac Surg. 2024;35(7):2194–2199. https://doi.org/10.1097/SCS.0000000000010386
[7] Yan L, Wang J, Cai X, et al. Macrophage plasticity: signaling pathways, tissue repair, and regeneration. MedComm (2020). 2024;5(8):e658. https://doi.org/10.1002/mco2.658
[8] Li X, Ren Y, Chang K, et al. Adipose tissue macrophages as potential targets for obesity and metabolic diseases. Front Immunol. 2023;14:1153915. https://doi.org/10.3389/fimmu.2023.1153915
[9] Dang J, Yang J, Yu Z, et al. Bone marrow mesenchymal stem cells enhance angiogenesis and promote fat retention in fat grafting via polarized macrophages. Stem Cell Res Ther. 2022;13(1):52. https://doi.org/10.1186/s13287-022-02709-2
[10] Li J, Guo T, Li Y, et al. Adipose stem cells regulate lipid metabolism by upregulating mitochondrial fatty acid β-oxidation in macrophages to improve the retention rate of transplanted fat. Stem Cell Res Ther. 2024;15(1):328. https://doi.org/10.1186/s13287-024-03953-4
[11] Bi X, Li Y, Dong Z, et al. Recent developments in extracellular matrix remodeling for fat grafting. Front Cell Dev Biol. 2021;9:767362. https://doi.org/10.3389/fcell.2021.767362
[12] Vania R, Sanjaya IG, Hamid AR, et al. Early activation of macrophage-2 with IL-4 in stromal vascular fraction increases VEGF levels and adipocyte count and maintains volume of fat graft in Wistar rats (Rattus norvegicus). Narra J. 2024;4(3):e1080. https://doi.org/10.52225/narra.v4i3.1080
[13] Lin J, Zhu S, Liao Y, et al. Spontaneous browning of white adipose tissue improves angiogenesis and reduces macrophage infiltration after fat grafting in mice. Front Cell Dev Biol. 2022;10:845158. https://doi.org/10.3389/fcell.2022.845158
[14] Wang J, Du H, Xie W, et al. CAR-macrophage therapy alleviates myocardial ischemia-reperfusion injury. Circ Res. 2024;135(12):1161–1174. https://doi.org/10.1161/CIRCRESAHA.124.325212
[15] Chen X, Chen Y, Wang Z, et al. Adipose-derived stem cells regulate CD4+ T-cell-mediated macrophage polarization and fibrosis in fat grafting in a mouse model. Heliyon. 2022;8(11):e11538. https://doi.org/10.1016/j.heliyon.2022.e11538
[16] Xue JD, Gao J, Tang AF, et al. Shaping the immune landscape: multidimensional environmental stimuli refine macrophage polarization and foster revolutionary approaches in tissue regeneration. Heliyon. 2024;10(17):e37192. https://doi.org/10.1016/j.heliyon.2024.e37192
[17] Zhao L, Tang S, Chen F, et al. Regulation of macrophage polarization by targeted metabolic reprogramming for the treatment of lupus nephritis. Mol Med. 2024;30(1):96. https://doi.org/10.1186/s10020-024-00866-z
[18] Wang P, Li Z, Song Y, et al. Resveratrol-driven macrophage polarization: unveiling mechanisms and therapeutic potential. Front Pharmacol. 2025;15:1516609. https://doi.org/10.3389/fphar.2024.1516609
[19] Klinkert K, Whelan D, Clover AJP, et al. Selective M2 macrophage depletion leads to prolonged inflammation in surgical wounds. Eur Surg Res. 2017;58(3–4):109–120. https://doi.org/10.1159/000451078
[20] Peng C, Feng Z, Ou L, et al. Syzygium aromaticum enhances innate immunity by triggering macrophage M1 polarization and alleviates helicobacter pylori-induced inflammation. J Funct Foods. 2023;107:13. https://doi.org/10.1016/j.jff.2023.105626
[21] Shen L, Li Y, Zhao H. Fibroblast growth factor signaling in macrophage polarization: impact on health and diseases. Front Immunol. 2024;15:1390453. https://doi.org/10.3389/fimmu.2024.1390453
[22] Chen J, Wang Y, Tang T, et al. Correction: enhanced effects of slowly co-released TGF-β3 and BMP-2 from biomimetic calcium phosphate-coated silk fibroin scaffolds in the repair of osteochondral defects. J Nanobiotechnol. 2025;23(1):46. https://doi.org/10.1186/s12951-025-03131-5
[23] Peng F, Zhang X, Wang Y, et al. Guided bone regeneration in long-bone defect with a bilayer mineralized collagen membrane. Collagen Leather. 2023;5(1):36. https://doi.org/10.1186/s42825-023-00144-4
[24] Paskal W, Gotowiec M, Stachura A, et al. VEGF and other gene therapies improve flap survival – a systematic review and meta-analysis of preclinical studies. Int J Mol Sci. 2024;25(5):2622. https://doi.org/10.3390/ijms25052622
[25] Algahtani M, Natarajan U, Alhazzani K, et al. Evaluation of anti-angiogenic agent F16 for targeting glioblastoma xenograft tumors. Cancer Genet. 2022;264–265:71–89. https://doi.org/10.1016/j.cancergen.2022.04.001
[26] Li R, Huang Y, Liu H, et al. Abstract 2162: comparing volatile and intravenous anesthetics in a mouse model of breast cancer metastasis. Cancer Res. 2018;78(13 Suppl.):2162. https://doi.org/10.1158/1538-7445.AM2018-2162