Klaus Kirketerp-Møller1, Thomas Bjarnsholt2,3, Peter Østrup Jensen2–4 and Magnus S. Ågren1,5,6
1Copenhagen Wound Healing Center and 5Digestive Disease Center, Bispebjerg Hospital, Bispebjerg Bakke 23, DK-2400 Copenhagen NV, 2Costerton Biofilm Center, Department of Clinical Immunology and Microbiology, University of Copenhagen, 3Department of Clinical Microbiology, Rigshospitalet, 4Institute for Inflammation Research, Center for Rheumatology and Spine Diseases, Copenhagen University Hospital, Rigshospitalet, Copenhagen, Denmark, and 6Department of Clinical Medicine, Faculty of Health and Medical Sciences, University of Copenhagen, Copenhagen, Denmark. E-mail: kkm@dadlnet.dk
Accepted Jun 23, 2020; Epub ahead of print Jul 3, 2020
Acta Derm Venereol 2020; 100: adv00232.
Non-healing cutaneous wounds are commonly associated with dysregulation of the zinc-dependent matrix metalloproteinases (MMPs) (1). MMP-8 is the most efficient human collagenase and degrades collagens synergistically with MMP-9 (2, 3). Although MMP-8 and MMP-9 are necessary for normal wound healing, their excessive expression impairs experimental wound healing (3–6).
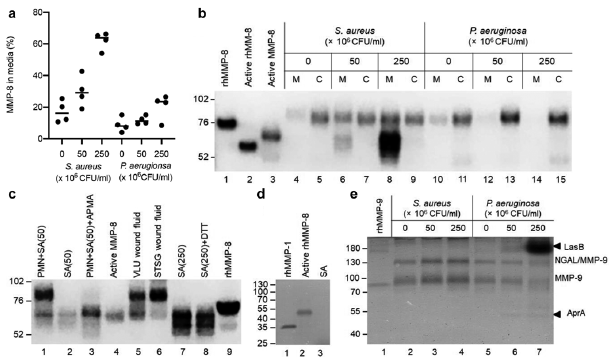
Elevated MMP-8 and MMP-9 levels were found in colonized venous leg ulcers (VLUs) compared with sterile split-thickness skin graft (STSG) donor sites in patients with VLUs. Polymorphonuclear leukocytes (PMNs) are the major sources of MMP-8 and MMP-9 in VLUs (7). These MMPs are pre-formed and stored in the secondary (8) and tertiary (9) granules of mature PMNs. It was hypothesized that the presence of bacteria results in a species-specific increase in MMP-8 and MMP-9 levels by stimulating degranulation of PMNs and promoting the conversion of catalytically inactive MMPs to their active forms via bacterial-derived proteinases. Staphylococcus aureus and Pseudomonas aeruginosa were studied here as prototypic wound pathogens (10).
Human PMNs were exposed to sterile-filtered supernatants from S. aureus and P. aeruginosa grown under static biofilm conditions. The PMN viability (> 70%) was similar in the control and treatment groups, and the mean ± standard deviation [SD] total MMP-8 content in 106 PMNs from the 4 donors was 39 ± 2.4 ng (8). S. aureus supernatants increased (p < 0.00050) the release of MMP-8 from the PMNs (Fig. 1a); the release with the highest titre was 62 ± 5.5%, which was higher than the middle titre (30 ± 9.9%, p = 0.0010) and the control (17 ± 6.9%, p < 0.00050). P. aeruginosa supernatants had a modest effect (p = 0.041) with an increased MMP-8 release with the highest titre (20 ± 8.1%) compared with the control (8.9 ± 4.6%, p = 0.040), but not compared with the middle titre (12 ± 2.5%, p = 0.75). The secretagogue phorbol 12-myristate 13-acetate (100 ng/ml) caused a release of 81 ± 2.6% compared with 3.8 ± 1.0% for the Krebs-Ringer balanced salt solution with 10 mM glucose control (8). No MMP-8 was detected in the bacterial supernatants with the ELISA assay. To ascertain whether the bacteria were capable of activating MMP-8, PMN media and cell lysates were analysed with Western blot. Prominent bands corresponding to active PMN-derived MMP-8 were observed in the media from S. aureus-treated PMNs (Fig. 1b, c) and in VLU wound fluids (lane 5, Fig. 1c). In contrast, minimal active MMP-8 was observed in sterile STSG donor site wound fluids (lane 6, Fig. 1c). S. aureus alone produced a protein (~65 kDa) that was recognized by the monoclonal antibody at the position of human active PMN-derived MMP-8 and remained unaltered by reduction (lanes 2, 7 and 8, Fig. 1c). Staphylococcus aureus strains can make 7 serine proteinases (SspA and SplA-F), 2 cysteine proteinases (ScpA and SspB), and a single metalloproteinase (Aur) (13). This protein was unlikely to be a metalloproteinase (Fig. 1d). Thus, we were unable to conclude whether S. aureus products activate latent PMN-derived MMP-8.
In accordance with the MMP-8 findings, S. aureus products stimulated the exocytosis of MMP-9, while P. aeruginosa mitigated the MMP-9 levels in media. Judged by the zymographic analysis, MMP-9 released from the PMNs was latent. The 2 gelatinolytic bands only observed in lanes 6 and 7 in Fig. 1e presumably correspond to the action of LasB complex and AprA metalloproteinases produced by P. aeruginosa (14).
This study demonstrated that bacterial products exert a major impact on the release of MMP-8 and MMP-9 from immune cells present in wounds. These effects appear to be species-dependent, and S. aureus biofilm supernatants increased MMP-8 and MMP-9 release from human PMNs, while P. aeruginosa appeared to decrease the medium levels of these MMPs. This study was unable to explain the discrepancy between the ELISA and the Western blot measurements of MMP-8. Nevertheless, the increased degranulation of PMNs may contribute to the elevated MMP-8 and MMP-9 levels in wounds with bacterial biofilms.
For our initial important findings to be translated to the microenvironment of wounds, they should be expanded by including a wide range of different clinical isolates. Furthermore, because the wound microbiota is commonly polymicrobial, the combined effect of different bacterial cell types should be evaluated (10). In these experiments, the effects of products produced by bacteria grown under various conditions and modes (planktonic growth vs biofilm formation) ought to be assessed. Supernatants from PAO1 grown in a TSB medium with agitation activated latent MMP-9 produced by epithelial cells (15).
The results of this study suggest that the interpatient variations in MMP-8 and MMP-9 levels and the degree of activation can, at least in part, be ascribed to differences in the microbiota of wounds.
The authors thank Omid Niazi, Vibeke Pless, Anne Kirstine Nielsen and Marjan Yousefi for their expert technical assistance.
The authors have no conflicts of interest to declare.