ORIGINAL REPORT
Higher Expression of Lympho-epithelial Kazal-type-Related Inhibitor-1 Fragments and Decreased Desquamation in the Lesional Skin of Nummular Eczema
Sara Estefania MONTENEGRO1-3#, Jang-Hee OH1-3#, Joong Heon SUH1-3, Je-Ho MUN1-3 and Jin Ho CHUNG1-4
1Department of Dermatology, Seoul National University College of Medicine, Seoul, Republic of Korea, 2Institute of Human-Environment Interface Biology, Medical Research Center, Seoul National University, Seoul, Republic of Korea, 3Laboratory of Cutaneous Aging Research, Biomedical Research Institute, Seoul National University Hospital, Seoul, Republic of Korea and 4Institute on Aging, Seoul National University, Seoul, Republic of Korea
#These authors contributed equally to this work.
Nummular eczema, a chronic dermatitis characterized by coin-shaped lesions, was first documented in 1857. However, its pathophysiological characteristics are still not well known. To investigate differences in the regulation of the desquamation process in the stratum corneum of lesional and nonlesional skin of patients with nummular eczema and healthy control subjects, tape-stripped stratum corneum samples from patients with nummular eczema and healthy volunteers were analysed using immunofluorescence staining and western blot analysis. In the nummular eczema lesional skin, expression of desmoglein-1, desmocollin-1, and corneodesmosin exhibited a disorganized, dense or partially diffuse non-peripheral pattern with increased intensity, compared with the peripheral patterns observed in healthy or nonlesional skin, suggesting the impaired desquamation process in nummular eczema. Furthermore, although the expression of the desquamation-related serine proteases, kallikrein-related peptidase 7 and 5, was increased in nummular eczema lesional skin, the immunofluorescence staining of lympho-epithelial Kazal-type-related inhibitor-1, an endogenous inhibitor of various kallikrein-related peptidases, and its fragments were significantly increased in the nummular eczema lesional skin, suggesting its contribution to the inhibition of corneodesmosomal degradation. Therefore, the increased detection of corneodesmosomal proteins in nummular eczema lesions may be due to the increased amount of the fragments of lympho-epithelial Kazal-type-related inhibitor-1, which could contribute to delayed desquamation.
Key words: corneodesmosin; desmocollin-1; desmoglein-1; lympho-epithelial Kazal-type-related inhibitor; nummular eczema.
SIGNIFICANCE
Nummular eczema is clinically characterized by coin-shaped or oval pruritic lesions with well-defined borders, usually on the legs, but also on other areas. Its precise underlying cause is not yet fully understood. We discovered an elevated expression of corneodesmosomal proteins in the lesional skin of patients with nummular eczema. This increase is potentially mediated by an elevation in lympho-epithelial Kazal-type-related inhibitor-1, which inhibits the proteases involved in desquamation. This inhibition may lead to delayed desquamation and the emergence of scales. Therefore, we suggest that antagonists targeting lympho-epithelial Kazal-type-related inhibitor-1 may serve as potential therapeutic candidates for the treatment of nummular eczema.
Citation: Acta Derm Venereol 2024; 104: 18636. DOI https://doi.org/10.2340/actadv.v104.18636.
Copyright: © Published by Medical Journals Sweden, on behalf of the Society for Publication of Acta Dermato-Venereologica. This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial 4.0 International License (https://creativecommons.org/licenses/by-nc/4.0/)
Submitted: Sep 3, 2023; Accepted: Mar 4, 2024; Published: Mar 29, 2024
Corr: Jin Ho Chung, MD, PhD, Department of Dermatology, Seoul National University College of Medicine, 101 Daehak-ro, Jongno-gu, Seoul, 03080, Republic of Korea. E-mail: jhchung@snu.ac.kr
Competing interests and funding: The authors have no conflicts of interest to declare.
INTRODUCTION
Nummular eczema (NE) is clinically characterized by coin-shaped or oval pruritic lesions with well-defined borders, usually on the legs, but also on other areas, including the arms and trunk (1–3). NE lesions typically exhibit papules/vesicles (acute phase) and crusts/desquamation (chronic phase) (4). The exact cause of NE is unknown. However, several factors including nutrition, infection, excessive alcohol intake, and dry skin, particularly in the elderly, have been linked to its development (5–9).
The formation of scales is a common feature of xerosis and various eczemas, including NE, and is caused by an imbalance of various proteases and endogenous inhibitors (4, 10–12). In dry skin or atopic dermatitis (AD) skin, increased non-peripheral corneodesmosome structures in the upper stratum corneum (SC) with an increase in corneodesmosomal proteins such as corneodesmosin (CDSN), desmoglein-1 (DSG1), and desmocollin-1 (DSC1) have been reported (12–14), suggesting the impaired desquamation process.
Corneodesmosomes, the modified desmosomes of the uppermost denucleated epidermal layers, are composed of DSG1, DSC1, and CDSN (15). DSG1 and DSC1 are desmosomal cadherins, expressed in the superficial layers of the epidermis, and mediate adhesion in desmosomes and corneodesmosomes (16, 17). CDSN is a 52–56 kDa glycoprotein expressed in cornified epithelia and the inner root sheath of hair follicles (18, 19). In the epidermis, CDSN is secreted into the SC through lamellar bodies, and reinforces corneodesmosomes in the SC (20, 21).
Corneodesmosomes in the upper SC are normally degraded by proteases such as kallikrein-related peptidases (KLKs) during desquamation (15, 18). KLK7 mediates the disruption of corneodesmosomes by hydrolysing the CDSN and DSC1, but not DSG1, while KLK5 cleaves all three components (22, 23). KLK1, 6, and 14 can also degrade DSG1 (24). Epidermal KLK activities are strictly controlled by endogenous specific inhibitors, including lympho-epithelial Kazal-type-related inhibitor (LEKTI)-1 (23, 25). LEKTI-1 is processed into single or multiple domains and inhibits various proteinases, including epidermal KLK5, 6, 7, 13, and 14 (25, 26). Full-length or partial recombinant forms of LEKTI-1 are found to inhibit various proteases including trypsin, plasmin, chymotrypsin, and caspase-14, as well as KLKs (26, 27).
In this study, we aimed to better understand the pathophysiology of NE by investigating changes in the expression of corneodesmosomal components and their regulatory proteins, KLK5, 7, and LEKTI-1, in tape-stripped SC samples from NE skin and healthy skin.
MATERIALS AND METHODS
Study/ethical approval
The protocol for obtaining skin tissues from patients with NE and healthy volunteers using the tape-stripping method was approved by the medical ethics committee of the Institutional Review Board of Seoul National University Hospital (IRB No. H-1805-071-946), and all participants provided written informed consent. The study was conducted in accordance with the principles described in the Declaration of Helsinki. Patients with NE who visited Seoul National University Hospital between August 2018 and January 2019 and aged-matched healthy volunteers were involved in this study. Diagnosis of NE was performed by a board-certified dermatologist based on clinical evaluation, including using eczema area and severity index (EASI) (28) and investigator’s global assessment (IGA; 0 to 5 points) scores. Patient characteristics are summarized in Table SI. Patients with NE who had other acute diseases or who were administered anti-inflammatory medicines for NE within 3 months were excluded.
Tape-stripping of stratum corneum
SC samples were collected from the lesional and nonlesional areas in the leg, arm, abdomen, or back skin of patients with NE and the same skin areas of healthy volunteers using Scotch® tape (3M, St Paul, MN, USA). For western blot analysis, the 1.8 cm × 5 cm Scotch® tape was placed on the lesional and nonlesional skin of patients with NE (mean age ± standard error of the mean (SEM), 51.3 ± 2.72 years; age range, 27–71 years; 12 females and 12 males) or skin of healthy volunteers (48.9 ± 2.12 years; age range, 36–59 years; 10 females and 5 males) for 3 minutes, applying pressure during the last minute, and then attached and detached 10 times. For immunofluorescence staining, the SC samples were obtained using the 0.84 cm × 3 cm double-sided adhesive tape (3M) on the slide by placing the tape-adhered slide on the skin of patients with NE or healthy volunteers for 3 minutes, applying pressure during the last minute. Tapes and slides containing SC tissues were stored at –20°C until use.
Protein extraction from stripped tapes and western blot analysis
The stripped tapes were cut into smaller pieces, placed into microtubes, and 500 μL of modified 6% SDS sample buffer (20% glycerol, 6% SDS, 125 mM Tris-HCl, pH 6.8) was added to the samples, and boiled at 95 °C for 5 min. Thereafter, supernatants were collected by centrifugation at 13,000 rpm at 4°C for 5 min, and the protein concentration was measured using a bicinchoninic acid assay (Sigma-Aldrich, St Louis, MO, USA). For western blot analysis, the same amounts of protein were added to 5% β-mercaptoethanol and 0.02% bromophenol blue (Sigma-Aldrich), separated using SDS-PAGE on 8–15% gels, and transferred to a nitrocellulose membrane (GE Healthcare Life Science, Chicago, IL, USA). Then, membranes were stained with Ponceau S solution (Elpis Biotech, Daejeon, Korea) and blocked with 5% skim milk in Tris-buffered saline containing 0.1% Tween® 20 (TBS-T), and incubated with primary antibodies diluted in TBS-T overnight at 4°C. Information for the primary antibodies is summarized in Table SII. Subsequently, the membranes were incubated with horseradish peroxidase-conjugated anti-mouse, rabbit, or rat IgG secondary antibodies (GeneTex, Irvine, CA, USA). Chemiluminescence signals were obtained using ECL reagents (Thermo Fisher Scientific, Waltham, MA, USA). The intensity of each band was normalized to the intensity of Ponceau S-stained bands between 50 and 70 kDa, using ImageJ software (National Institutes of Health, Bethesda, MD, USA).
Immunofluorescence staining
The SC samples on slides were blocked using an UltraVision protein block (Thermo Fisher Scientific), washed with PBS (1×), and incubated with primary antibodies overnight at 4°C. Information for the primary antibodies is summarized in Table SII. The slides were then washed with PBS, and incubated with secondary antibodies: anti-mouse, anti-rabbit, or anti-rat IgG conjugated with Alexa 594® or Alexa 488® (1:400, Thermo Fisher Scientific). The slides were mounted on Shandon Immu-Mount (Thermo Fisher Scientific). Fluorescent images were obtained using an Eclipse Ni microscope with a DS-Ri2 camera (Nikon, Tokyo, Japan), under identical conditions for each target protein, except the weak signal images of the lesional samples, which were obtained by adjusting the exposure time to show the patterns clearly. The intensity of immunofluorescence signal was analysed using ImageJ.
Statistical analysis
Statistical analyses were performed using IBM SPSS Statistics 25 (IBM Corp, Armonk, NY, USA). The Wilcoxon signed-rank test was used to compare paired samples of lesional and nonlesional skin of the same patients with NE, while the Mann–Whitney U test was used for non-paired samples of healthy skin and lesional or nonlesional skin from patients with NE. Statistical significance was set at p < 0.05.
RESULTS
Higher corneodesmosomal protein expression in the lesional skin of patients with nummular eczema
We first investigated the corneodesmosomal protein expression in the tape-stripped uppermost layers of the SC of the lesional and nonlesional skin from patients with NE and healthy skin from normal subjects by performing immunofluorescence staining for the corneodesmosomal structural proteins, DSG1, DSC1, and CDSN. Immunofluorescence staining of DSG1, DSC1, and CDSN in the SC from healthy skin showed a peripheral distribution, as reported in previous studies (13,29). In contrast, in lesional skin of patients with NE, DSG1, DSC1, and CDSN exhibited a disorganized, dense, or partially diffuse non-peripheral pattern with increased intensity (Fig. 1). Meanwhile, nonlesional skin of NE exhibited a predominantly peripheral distribution pattern mixed with a partial diffuse pattern with a partial increase in intensity for DSG1, DSC1, and CDSN (Fig. 1). These results suggest that the desquamation process may be impaired in lesional skin of patients with NE.
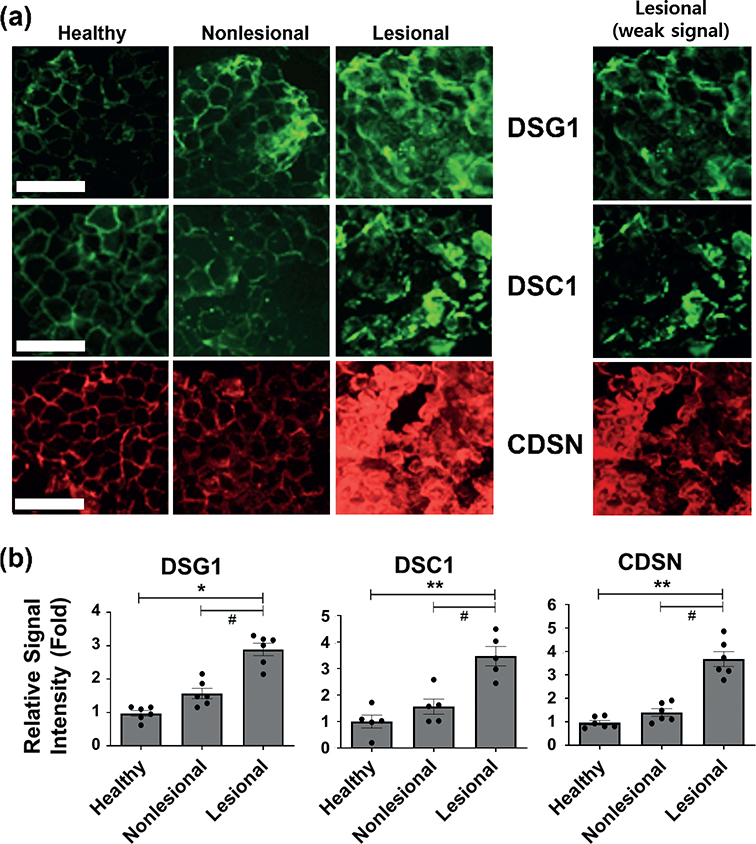
Fig. 1. Higher corneodesmosomal protein expression using immunofluorescence staining in patients with nummular eczema (NE). (a) Immunofluorescence staining images of desmoglein-1 (DSG1), desmocollin-1 (DSC1), and corneodesmosin (CDSN) in tape-stripped uppermost stratum corneum (SC) of healthy subjects and nonlesional and lesional SC of the same patient with nummular eczema (NE), n = 5 or 6 for each group. Scale bar = 100 μm. The same images of the lesional SC with weak signal were additionally shown for clearer pattern of staining in the lesional SC samples. (b) Relative signal intensity of immunofluorescence staining was quantified and represented as mean fold ± SEM. #p < 0.05; lesional skin vs nonlesional skin of the same patients with NE (Wilcoxon signed-rank test); *p < 0.05, **p < 0.01 lesional or nonlesional skin vs the healthy skin (Mann–Whitney U test).
To confirm those changes, we performed western blot analyses for these proteins with tape-stripped SC samples from the patients with NE and healthy volunteers. In previous studies, DSG1 intact forms were detected around 150–160 kDa, and its processed forms were mainly detected at 80–95 kDa in tape-stripped SC samples from normal and winter xerosis skin (12,30). In this study, healthy SC showed mainly fragmented forms of DSG1 (50–70 kDa). On the other hand, NE skin showed stronger signals of both fragmented forms (50–100 kDa) and intact form (~150 kDa) of DSG1, along with smaller-sized fragments (< 50 kDa) in almost all samples (Fig. 2a). Ponceau S stain bands (50–70 kDa) were used as a loading control. The densitometry of the western blot analysis of 15 healthy and 18 NE SC samples revealed that the expression of DSG1 was significantly higher in lesional skin of patients with NE (intact forms+fragmented forms) than in nonlesional skin of the same patients. Furthermore, DSG1 expression was also significantly higher in both lesional and nonlesional skin of patients with NE than in healthy controls (Fig. 2b).
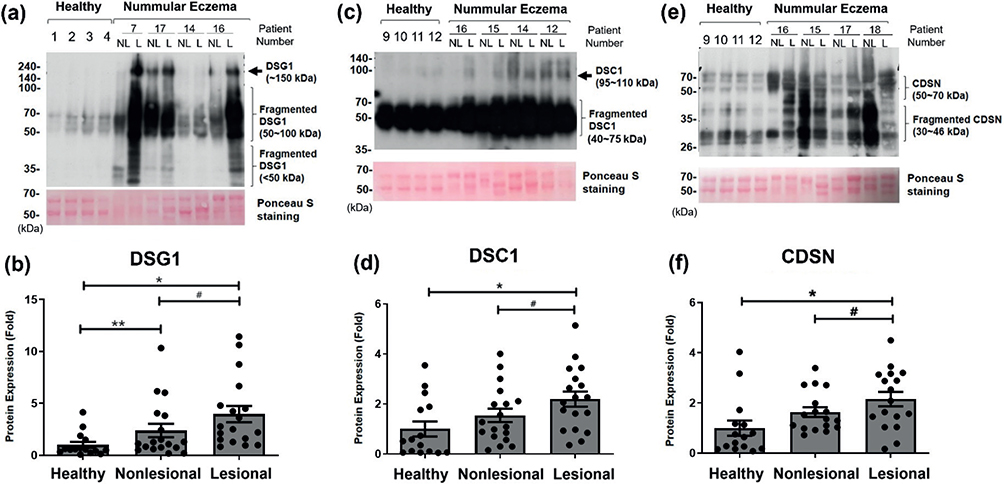
Fig. 2. Higher corneodesmosomal protein expression using western blot analysis in patients with nummular eczema (NE). Protein expressions of (a, b) DSG1, (c, d) DSC1, and (e, f) CDSN in tape-stripped SC samples from healthy skin, or nonlesional (NL) and lesional (L) skin of the same patient with NE were analysed using western blot. Ponceau S stain was used as loading control. Results of each 4 representatives of healthy and NE samples are shown. Each band of (b) DSG1, (d) DSC1, and (f) CDSN was analysed using densitometry, and the densitometric data (intact forms+fragmented forms) are presented as the mean fold ± SEM. Intensity of each band was normalized with the intensity of Ponceau S-stained bands between 50 and 70 kDa (patients with NE, n = 17 or 18; healthy skin, n = 15). #p < 0.05; lesional skin vs nonlesional skin of the same patients with NE (Wilcoxon signed-rank test); *p < 0.05, **p < 0.01 lesional or nonlesional skin vs the healthy skin (Mann–Whitney U test). Positive correlations between these corneodesmosomal protein levels were confirmed by the linear regression analysis (Fig. S2).
The intact form of DSC1 has been known to be 95–110 kDa in size (31,32), but the size of its fragmented form in SC has not been reported. In our results, healthy skin mainly showed fragmented forms of DSC1 (40–70 kDa), and almost all SC samples from NE lesional skin showed stronger signals of both fragmented and intact forms of DSC1 (Fig. 2c). Because of the detection of DSC1 intact forms, the fragmented forms were overexposed in Fig. 2c; therefore, we provide a distinguishable image for DSC1 fragmented forms in Fig. S1. The densitometry of 15 healthy and 18 NE SC samples revealed a significant increase in the expression of DSC1 (intact forms+fragmented forms) in lesional skin compared with nonlesional skin of the same patient and healthy skin (Fig. 2d).
The intact form of CDSN (52–56 kDa) has been reported to fragment into 40–48 or 33–36 kDa during SC maturation (19), and the fragmented forms were mainly detected in tape-stripped SC samples from normal and winter xerosis skin (12). The western blot analysis of the 15 healthy and 17 NE SC samples detected both intact (50–70 kDa) and fragmented forms (30–46 kDa) of CDSN, and showed a higher expression of CDSN (intact forms+fragmented forms) in lesional skin than nonlesional skin of the same patient and healthy skin (Fig. 2e and f). DSC1 and CDSN expression in nonlesional skin tends to be higher than in healthy skin, but not significant (Fig. 2d and f).
This increased expression and altered distribution of DSG1, DSC1, and CDSN in lesional skin of patients with NE suggests potential disruption in the process of corneodesmosomal protein degradation related to desquamation.
Higher KLK7 and KLK5 expression in the lesional skin of patients with nummular eczema
Because the degradation of corneodesmosomal protein in the normal skin is well known to be mediated by KLKs, such as KLK7 and KLK5 (23), we investigated the expression of KLK7 and KLK5.
The immunofluorescence staining showed that KLK7 and KLK5 had a peripheral distribution pattern in healthy and nonlesional skin (Fig. 3a), as previously reported (13). However, in lesional skin of patients with NE, KLK7 and KLK5 immunofluorescence staining exhibited a disorganized, dense, or partially diffuse non-peripheral pattern with increased intensity (Fig. 3a and b).
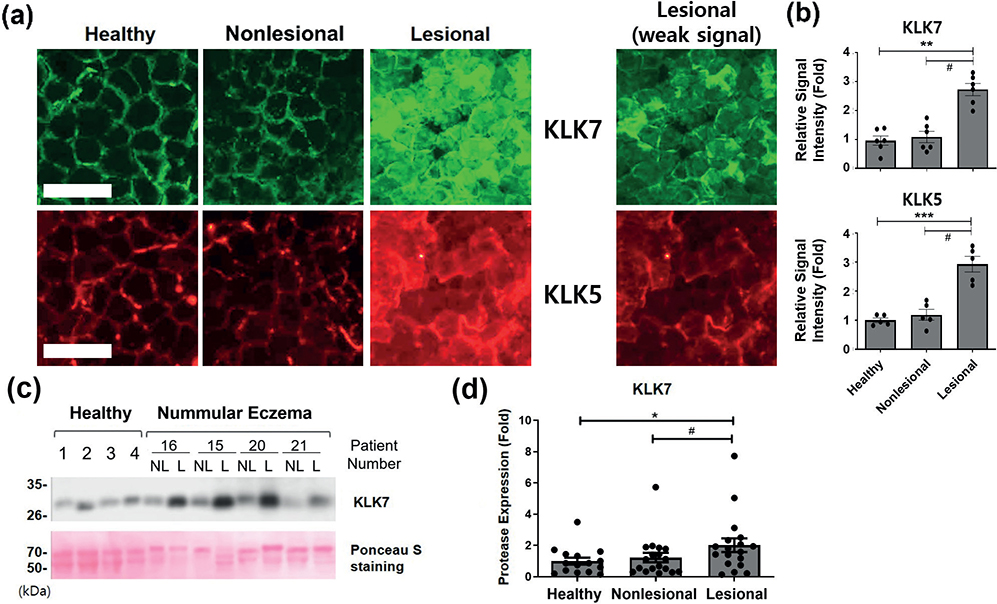
Fig. 3. Higher kallikrein-related peptidase (KLK) 7 and 5 expression in patients with nummular eczema (NE). (a) Immunofluorescence staining images of KLK7 and KLK5 in tape-stripped uppermost SC of healthy subject skin and nonlesional and lesional skin of the same patient with NE, n = 5 or 6 for each group. Scale bar = 100 μm. The same images of the lesional SC with weak signal were additionally shown for clearer pattern of staining in the lesional SC samples. (b) Relative signal intensity of immunofluorescence staining was quantified and represented as mean fold ± SEM. (c) Protein expressions of KLK7 in tape-stripped SC samples from healthy skin, or nonlesional (NL) and lesional (L) skin of the same patient with NE were analysed using western blot. Ponceau S stain was used as loading control. Results of each 4 representatives of healthy and NE samples are shown. (d) Each band of KLK7 was analysed using densitometry, and the densitometric data are presented as the mean fold ± SEM. Intensity of each band was normalized with the intensity of Ponceau S-stained bands between 50 and 70 kDa (patients with NE, n = 18; healthy skin, n = 15). #p < 0.05; lesional skin vs nonlesional skin of the same patients with NE (Wilcoxon signed-rank test); *p < 0.05, lesional or nonlesional skin vs the healthy skin (Mann–Whitney U test).
The western blot analyses of 15 healthy and 18 NE SC samples also revealed a significant increase in the protein level of KLK7 (between 26–35 kDa) in lesional skin of patients with NE, compared with nonlesional skin of the same patient or healthy skin, while there was no significant difference in KLK7 expression between nonlesional NE skin and healthy skin (Fig. 3c and d).
Therefore, to understand why corneodesmosomal proteins are increased in NE despite the increase in their degradation enzymes KLK7 and KLK5, the expression of LEKTI-1, an endogenous inhibitor of these proteinases, was further investigated.
Higher expression of LEKTI-1 in the lesional skin of patients with nummular eczema
Immunofluorescence staining of LEKTI-1 showed a peripheral but partly spot-like discontinuous distribution pattern in healthy skin and nonlesional skin; however, in lesional skin, immunofluorescence staining of LEKTI-1 also exhibited a disorganized and dense or partially diffuse non-peripheral pattern with strongly increased intensity (> 6 folds to healthy skin) (Fig. 4a and b).
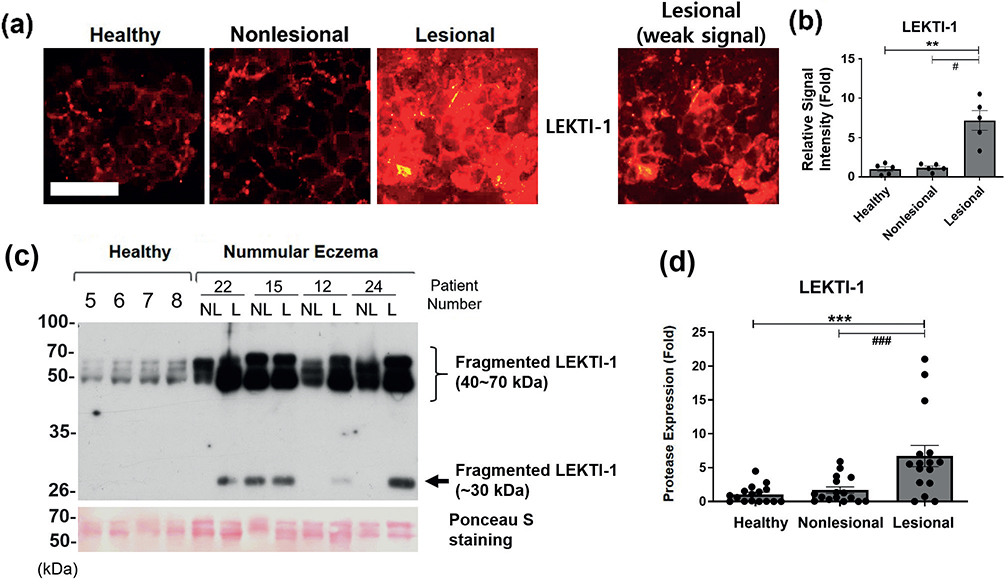
Fig. 4. Higher expression of lympho-epithelial Kazal-type-related inhibitor (LEKTI)-1 in patients with nummular eczema (NE). (a) Immunofluorescence staining images of LEKTI-1 in tape-stripped uppermost SC of healthy subject skin and nonlesional and lesional skin of the same patient with NE, n = 5 for each group. Scale bar=100 μm. The same image of the lesional SC with weak signal was additionally shown for clearer pattern of staining in the lesional SC sample. (b) Relative signal intensity of immunofluorescence staining was quantified and represented as mean fold ± SEM. (c) Protein expression of LEKTI-1 fragments in tape-stripped SC samples from healthy skin, NE nonlesional (NL), and lesional (L) skin were analysed using western blot. Ponceau S stain was used as loading control. Results of four representatives of healthy and NE samples are shown. (d) Each band of LEKTI-1 was analysed using densitometry, and the densitometric data (sum of all bands) are presented as the mean fold ± SEM. Intensity of each band was normalized with the intensity of Ponceau S-stained bands between 50 and 70 kDa (patients with NE, n = 12; healthy skin, n = 12). #p < 0.05; lesional skin vs nonlesional skin of the same patients with NE (Wilcoxon signed-rank test); *p < 0.05, lesional or nonlesional skin vs the healthy skin (Mann–Whitney U test).
In western blot, three precursor forms of LEKTI-1 (~125, ~145, and ~148 kDa) and active fragmented forms of various sizes (7–68 kDa) have been reported (26,33,34). Western blot analysis of 12 healthy and 12 NE SC samples showed a strongly increased detection of 40~70 kDa and ~30 kDa LEKTI-1 fragments in the NE lesional skin (> 6 folds to healthy skin), but weaker 40~70 kDa bands and no detectable ~30 kDa bands were observed in almost all nonlesional and healthy skin samples, except in a few cases of nonlesional skin (Fig. 4c and d). Therefore, from the results of immunofluorescence staining and western blot, the expression of LEKTI-1 in the SC of the lesional skin of patients with NE was increased, compared with healthy or nonlesional skin, and showed higher fold of increase than those of KLK7 and KLK5 (Fig. 4). Additionally, to determine actual activity of these proteases indirectly, we investigated serine protease activity by performing in situ casein zymography (13), and found that the net serine protease activity was almost negative in NE lesional skin, while healthy and NE nonlesional skin showed some extent of serine protease activity (Fig. S3), suggesting that KLK7 and KLK5 might be inhibited by increased LEKTI-1 in NE lesional skin.
These results suggest that the increased expression of LEKTI-1 fragments in lesional skin may inhibit desquamation-related proteases such as KLKs, leading to increased corneodesmosomal proteins and impaired desquamation in NE lesional skin.
DISCUSSION
To investigate and understand the molecular aspects of epidermal terminal differentiation in NE, we evaluated the changes in the corneodesmosomal components in the SC of patients with NE. Despite the increase in KLK7 and KLK5 expressions, an increase in levels and non-peripheral distribution of DSG1, DSC1, and CDSN was observed in the tape-stripped NE lesional skin SC, suggesting that the desquamation process is hindered in the NE lesional skin SC, and we found that the increased expression of LEKTI-1 in the lesional skin may play a role in inhibiting the desquamation process.
Increased amounts of corneodesmosomal proteins and non-peripheral distribution were found in the uppermost layers of SC in lesions of NE, which is similar to what is seen in dry skin or AD skin (12,13). Additionally, the detection of intact forms of corneodesmosomal proteins in the NE lesional samples also supports the idea that their degradation is partly inhibited, leading to the accumulation of these proteins and enhancing the adherence of the SC, contributing to the formation of scales in NE.
In AD skin, an increase in the expression of various KLKs was observed in SC (35, 36). However, the activities of trypsin-like or chymotrypsin-like proteases were not increased as much as expected by protein amount increases. In contrast, a much higher increase in furin-like protease activity was observed, which can activate LEKTI-1 (36). In addition, the impaired desquamation in AD is also found to be related to the incomplete activation of KLK7 and increased LEKTI-1 expression (13). Similarly, we also found increased immunofluorescence staining of LEKTI-1 and expression of LEKTI-1 fragments in the lesional skin of NE, more than the increases of KLK7 and KLK5.
LEKTI-1 is a multi-domain serine protease inhibitor protein, and its loss-of-function mutation causes Netherton syndrome (26,37). The precursor forms of LEKTI-1 are cleaved into smaller fragments in the post-endoplasmic reticulum compartment (26), and its variously sized active fragmented forms (7–68 kDa) have been found in conditioned medium of cultured epidermal keratinocytes or HEK293 cells transfected with LEKTI-1 isoforms, or in the extract of human epidermis (26,33,34). LEKTI-1 has 15 potential serine protease inhibitory domains (D1-D15), which have high homology with the typical Kazal-type serine protease inhibitors. However, 13 of these domains lack one of the three conserved disulphide bridges, with the exception of D2 and D15 (26). Based on the size of the detected LEKTI-1 fragments, they seem to include the previously reported 65/68 kDa, 42 kDa, and 31 kDa fragments of LEKTI-1 in human epidermis (26,34), which have potentials to inhibit KLK5, KLK7, or KLK14 (26,34). In addition, observation of no net serine protease activity in NE lesion skin also supports their possible inhibition by increased LEKTI-1 fragments. Thus, our findings suggest that the increase in the LEKTI-1 fragments in the NE lesional skin SC may contribute to the delayed desquamation, by leading to an imbalance in the interaction between corneodesmosomal protein-degrading proteases and their inhibitors. This results in erroneous corneodesmosomal assembly, enhancing adhesive strength and impaired desquamation.
In psoriasis, a higher level of LEKTI-1 expression is observed in the epidermis compared with AD or healthy skin, and TNFα, IL-17A, TLR1/2, 3, 5, and 2/6 ligands are known to induce the expression and secretion of LEKTI-1 in cultured epidermal keratinocytes (38,39). Therefore, the increased expression of LEKTI-1 in NE may also be a result of inflammatory cytokines- or TLR ligands-related responses.
Recently, it has been reported that NE has more mechanistic similarity to AD than psoriasis, though it has both type 2 and 3 immune signatures, and inhibition of type 2 immune response by treatment with dupilumab showed improvement of both type 2 and 3 immune signatures in patients with NE (40). This may support the similarity in increased LEKTI-1-mediated impairment of desquamation between NE and AD.
However, due to non-availability of skin tissue samples from patients with NE, we were unable to analyse the differences in the mRNA expression of these proteins. Thus, we could not verify the additional possibility that transcriptional upregulation of corneodesmosomal proteins may also contribute to the impairment of desquamation and disorganization of the SC. Furthermore, other mechanisms including incomplete activation of KLKs or their impaired secretion from lamellar granules, as reported in AD (13), were not investigated in this study. These are limitations of this study.
In conclusion, this study demonstrates that SC desquamation is impaired in the lesional skin of patients with NE. In particular, the increase in the LEKTI-1 fragments in the NE lesional skin suggests its contribution to the inhibition of desquamation-related proteinases like KLKs and the increase in the corneodesmosomal proteins, resulting in delayed skin desquamation.
ACKNOWLEDGEMENTS
This research was supported by a grant from the Korea Health Technology R&D Project through the Korea Health Industry Development Institute (KHIDI), funded by the Ministry of Health & Welfare, Republic of Korea (grant number: HI14C1277). The funders had no role in the design and conduct of the study; collection, management, analysis, and interpretation of the data; preparation, review, or approval of the manuscript; or decision to submit the manuscript for publication.
This study protocol was reviewed and approved by the medical ethics committee of the Institutional Review Board of Seoul National University Hospital (approval number: IRB No. H-1805-071-946), and all participants provided written informed consent.
REFERENCES
- Bonamonte D, Foti C, Vestita M, Ranieri LD, Angelini G. Nummular eczema and contact allergy: a retrospective study. Dermatitis 2012; 23: 153–157.
- Reich D, Psomadakis CE, Buka B. Nummular wczema. In: Reich D, Psomadakis CE, Buka B, editors. Top 50 dermatology case studies for primary care. Cham: Springer International Publishing; 2017: p. 167–172.
- Halberg M. Nummular eczema. J Emerg Med 2012; 43: e327–328.
- Lugovic-Mihic L, Bukvic I, Bulat V, Japundzic I. Factors contributing to chronic urticaria/angioedema and nummular eczema resolution: which findings are crucial? Acta Clin Croat 2019; 58: 595–603.
- Krogh HK. Nummular eczema. Its relationship to internal foci of infection. A survey of 84 case records. Acta Derm Venereol 1960; 40: 114–126.
- Tanaka T, Satoh T, Yokozeki H. Dental infection associated with nummular eczema as an overlooked focal infection. J Dermatol 2009; 36: 462–465.
- Aoyama H, Tanaka M, Hara M, Tabata N, Tagami H. Nummular eczema: an addition of senile xerosis and unique cutaneous reactivities to environmental aeroallergens. Dermatology 1999; 199: 135–139.
- Higgins EM, du Vivier AW. Cutaneous disease and alcohol misuse. Br Med Bull 1994; 50: 85–98.
- Jiamton S, Tangjaturonrusamee C, Kulthanan K. Clinical features and aggravating factors in nummular eczema in Thais. Asian Pac J Allergy Immunol 2013; 31: 36–42.
- Meyer-Hoffert U. Reddish, scaly, and itchy: how proteases and their inhibitors contribute to inflammatory skin diseases. Arch Immunol Ther Exp (Warsz) 2009; 57: 345–354.
- Nemeth V, Evans J. Eczema. In: StatPearls [Internet]. Treasure Island (FL): StatPearls Publishing; 2024 Jan. [cited 2022 Aug 8] Available from: https://www.ncbi.nlm.nih.gov/books/NBK538209/
- Simon M, Bernard D, Minondo AM, Camus C, Fiat F, Corcuff P, et al. Persistence of both peripheral and non-peripheral corneodesmosomes in the upper stratum corneum of winter xerosis skin versus only peripheral in normal skin. J Invest Dermatol 2001; 116: 23–30.
- Igawa S, Kishibe M, Minami-Hori M, Honma M, Tsujimura H, Ishikawa J, et al. Incomplete KLK7 secretion and upregulated LEKTI expression underlie hyperkeratotic stratum corneum in atopic dermatitis. J Invest Dermatol 2017; 137: 449–456.
- Delattre C, Winstall E, Lessard C, Donovan M, Simonetti L, Minondo AM, et al. Proteomic analysis identifies new biomarkers for postmenopausal and dry skin. Exp Dermatol 2012; 21: 205–210.
- Haftek M. Epidermal barrier disorders and corneodesmosome defects. Cell Tissue Res 2015; 360: 483–490.
- Whittock NV, Bower C. Targetting of desmoglein 1 in inherited and acquired skin diseases. Clin Exp Dermatol 2003; 28: 410–415.
- Kitajima Y. Regulation and impairments of dynamic desmosome and corneodesmosome remodeling. Eur J Dermatol 2013; 10.1684/ejd.2013.1976.
- Simon M, Jonca N, Guerrin M, Haftek M, Bernard D, Caubet C, et al. Refined characterization of corneodesmosin proteolysis during terminal differentiation of human epidermis and its relationship to desquamation. J Biol Chem 2001; 276: 20292–20299.
- Simon M, Montezin M, Guerrin M, Durieux JJ, Serre G. Characterization and purification of human corneodesmosin, an epidermal basic glycoprotein associated with corneocyte-specific modified desmosomes. J Biol Chem 1997; 272: 31770–31776.
- Richters RJH, Uzunbajakava NE, Timofeeva N, van de Kerkhof PCM, van Erp PEJ. Development of a novel approach to studying corneodesmosomes and stratum corneum adhesion: extending knowledge on the pathophysiology of sensitive skin. Skin Pharmacol Physiol 2019; 32: 81–93.
- Matsui T, Amagai M. Dissecting the formation, structure and barrier function of the stratum corneum. Int Immunol 2015; 27: 269–280.
- Fernandez IS, Standker L, Magert HJ, Forssmann WG, Gimenez-Gallego G, Romero A. Crystal structure of human epidermal kallikrein 7 (hK7) synthesized directly in its native state in E. coli: insights into the atomic basis of its inhibition by LEKTI domain 6 (LD6). J Mol Biol 2008; 377: 1488–1497.
- Caubet C, Jonca N, Brattsand M, Guerrin M, Bernard D, Schmidt R, et al. Degradation of corneodesmosome proteins by two serine proteases of the kallikrein family, SCTE/KLK5/hK5 and SCCE/KLK7/hK7. J Invest Dermatol 2004; 122: 1235–1244.
- Borgono CA, Michael IP, Komatsu N, Jayakumar A, Kapadia R, Clayman GL, et al. A potential role for multiple tissue kallikrein serine proteases in epidermal desquamation. J Biol Chem 2007; 282: 3640–3652.
- Ishida-Yamamoto A, Igawa S, Kishibe M, Honma M. Clinical and molecular implications of structural changes to desmosomes and corneodesmosomes. J Dermatol 2018; 45: 385–389.
- Deraison C, Bonnart C, Lopez F, Besson C, Robinson R, Jayakumar A, et al. LEKTI fragments specifically inhibit KLK5, KLK7, and KLK14 and control desquamation through a pH-dependent interaction. Mol Biol Cell 2007; 18: 3607–3619.
- Bennett K, Callard R, Heywood W, Harper J, Jayakumar A, Clayman GL, et al. New role for LEKTI in skin barrier formation: label-free quantitative proteomic identification of caspase 14 as a novel target for the protease inhibitor LEKTI. J Proteome Res 2010; 9: 4289–4294.
- Hanifin JM, Baghoomian W, Grinich E, Leshem YA, Jacobson M, Simpson EL. The eczema area and severity index: a practical guide. Dermatitis 2022; 33: 187–192.
- Lin TK, Crumrine D, Ackerman LD, Santiago JL, Roelandt T, Uchida Y, et al. Cellular changes that accompany shedding of human corneocytes. J Invest Dermatol 2012; 132: 2430–2439.
- Iwatsuki K, Harada H, Yokote R, Kaneko F. Differences in the expression of pemphigus antigens during epidermal differentiation. Br J Dermatol 1995; 133: 209–216.
- Delva E, Tucker DK, Kowalczyk AP. The desmosome. Cold Spring Harb Perspect Biol 2009; 1: a002543.
- Kopp T, Sitaru C, Pieczkowski F, Schneeberger A, Fodinger D, Zillikens D, et al. IgA pemphigus: occurrence of anti-desmocollin 1 and anti-desmoglein 1 antibody reactivity in an individual patient. J Dtsch Dermatol Ges 2006; 4: 1045–1050.
- Tartaglia-Polcini A, Bonnart C, Micheloni A, Cianfarani F, Andre A, Zambruno G, et al. SPINK5, the defective gene in netherton syndrome, encodes multiple LEKTI isoforms derived from alternative pre-mRNA processing. J Invest Dermatol 2006; 126: 315–324.
- Fortugno P, Bresciani A, Paolini C, Pazzagli C, El Hachem M, D’Alessio M, et al. Proteolytic activation cascade of the Netherton syndrome-defective protein, LEKTI, in the epidermis: implications for skin homeostasis. J Invest Dermatol 2011; 131: 2223–2232.
- Chavarria-Smith J, Chiu CPC, Jackman JK, Yin J, Zhang J, Hackney JA, et al. Dual antibody inhibition of KLK5 and KLK7 for Netherton syndrome and atopic dermatitis. Sci Transl Med 2022; 14: eabp9159.
- Komatsu N, Saijoh K, Kuk C, Liu AC, Khan S, Shirasaki F, et al. Human tissue kallikrein expression in the stratum corneum and serum of atopic dermatitis patients. Exp Dermatol 2007; 16: 513–519.
- Bitoun E, Micheloni A, Lamant L, Bonnart C, Tartaglia-Polcini A, Cobbold C, et al. LEKTI proteolytic processing in human primary keratinocytes, tissue distribution and defective expression in Netherton syndrome. Hum Mol Genet 2003; 12: 2417–2430.
- Sugihara S, Sugimoto S, Tachibana K, Kobashi M, Nomura H, Miyake T, et al. TNF-alpha and IL-17A induce the expression of lympho-epithelial Kazal-type inhibitor in epidermal keratinocytes. J Dermatol Sci 2019; 96: 26–32.
- Sugimoto S, Morizane S, Nomura H, Kobashi M, Sugihara S, Iwatsuki K. Toll-like receptor signaling induces the expression of lympho-epithelial Kazal-type inhibitor in epidermal keratinocytes. J Dermatol Sci 2018; 92: 181–187.
- Bohner A, Jargosch M, Muller NS, Garzorz-Stark N, Pilz C, Lauffer F, et al. The neglected twIn: nummular eczema is a variant of atopic dermatitis with codominant T(H)2/T(H)17 immune response. J Allergy Clin Immunol 2023; 152: 408–419.