SHORT COMMUNICATION
Topical Minoxidil Rejuvenates Hair Follicles from Men with Androgenetic Alopecia in Vivo
Assaf A. ZELTZER1, Aviad KEREN2, Ralf PAUS3,4 and Amos GILHAR2*
1Plastic & Reconstructive Surgery Department, Rambam Health Care Campus, Haifa, Israel, 2Skin Research Laboratory, Rappaport Faculty of Medicine, Technion – Israel Institute of Technology, Haifa, Israel, 3Dr. Phillip Frost Department of Dermatology & Cutaneous Surgery, Miller School of Medicine, University of Miami, Miami, FL, USA, and 4CUTANEON, Hamburg & Berlin, Germany. *E-mail: doritg2000@gmail.com
Citation: Acta Derm Venereol 2024; 104: adv24213. DOI https://doi.org/10.2340/actadv.v104.24213.
Copyright: © 2024 The Author(s). Published by MJS Publishing, on behalf of the Society for Publication of Acta Dermato-Venereologica. This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial 4.0 International License (https://creativecommons.org/licenses/by-nc/4.0/).
Submitted: Oct 24, 2023; Accepted: May 16, 2024; Published: Jun 11, 2024
Competing interests and funding: The authors state no conflict of interest. For the record, AG performs contract preclinical hair research for competing industry clients, including Johnson & Johnson, while RP is CEO of CUTANEON, a company that develops novel hair therapeutics.
This study was supported by the Technion Research and Development Foundation (TRDF) to A.G., and R.P. is a Frost Endowed Investigator in the Department of Dermatology, University of Miami.
INTRODUCTION
Male pattern androgenetic alopecia (mpAGA) is a progressive androgen-dependent hair loss disorder that affects androgen-sensitive hair follicles (HFs) in genetically predisposed individuals (1). One of the factors that may contribute to mpAGA development is oxidative stress, caused by imbalance between the generation and scavenging of reactive oxygen species (ROS), which may lead to decreased mitochondrial function (2), accelerate scalp skin ageing and may constitute an important link between skin ageing and AGA (3). In healthy human HFs, oxidative stress responses are tightly controlled, e.g., by NRF-2, or nuclear factor erythroid 2-related factor 2, which serves as a “master regulator” of cellular antioxidant response, activating genes that combat oxidative stress and protect cellular integrity (4), ROS scavenging enzymes, and intrafollicular production of melatonin (4). Moreover, dermal papilla cells obtained from AGA HFs reportedly show a down-regulation of vasculature-related genes (5). Given that vascular endothelial growth factor (VEGF) production by HF keratinocytes can control HF size and growth in mice (6), balding scalp HFs might have a diminished capacity to promote HF vascularization (5). The latter may aggravate an oxidative stress-accelerated skin and HF ageing phenotype in mpAGA skin, because the core angiogenesis-promoting growth factor, VEGF-A, is a key driver of human skin rejuvenation in vivo (7).
Taken together, this leads us to ask whether minoxidil might exert previously undetected anti-ageing effects on human HFs, namely at the level of oxidative damage responses, VEGF-A production, and/or mitochondrial function. This was further encouraged by evidence that minoxidil can stimulate VEGF production (8), and by our prior finding that hair growth inhibition (premature catagen) induced by perceived stress is significantly reduced after topical minoxidil application in mice in vivo (9), along with reduced degranulation of perifollicular mast cells – a major source of ROS generation around human HFs (10). Therefore, we probed whether topical minoxidil exerts anti-ageing effects in human mpAGA skin in vivo, i.e., our established humanized mpAGA mouse model (11, 12).
METHODS AND RESULTS
For this purpose, we re-analysed previously published mpAGA scalp skin fragments from men who had been xenotransplanted onto immunocompromised SCID/beige mice (11) and treated with topical minoxidil (5%) in vivo for changes in selected key ageing biomarkers, key mitochondrial read-outs, and VEGF-A protein expression (7). Lesional biopsies from frontal areas of 9 mpAGA patients (mean age: 35.9 ± 9.4 years) were obtained with written informed consent and Institutional Review Board (IRB) approval, and transplanted onto 9 SCID/beige mice. For 4 months, we administered 5% topical minoxidil daily to 5 mice and only the vehicle to another group of 4 mice, targeting the xenotransplants. Horizontal sections of the xenotransplants before and after transplantation were processed and immunostained as described previously (7) and are detailed in Table SI. This enabled a comprehensive analysis involving antibodies against lamin B1 (13), p16INK4A (14), protein S6 (p-S6) (15), collagen XVIIA (16), and biomarkers of mitochondrial activity and/or ageing (MTCO1, PGC1α, SIRT1, and porin/VDAC ), and both human and murine VEGF-A (7). Finally, we also interrogated oxidative damage responses by immunostaining NRF2 and some of its key downstream targets. Immunostaining results were analysed by quantitative immunohistomorphometry (QIHM).
Protein expression changes in response to the treatment were predominantly observed in the hair bulb region of the HFs (Table SII). QIHM revealed a significant upregulation in the protein expression of lamin B1 (p < 0.05) (13), a crucial marker associated with cellular structure and integrity, indicating an enhancement in cellular robustness and structural stability. Additionally, SIRT1 or sirtuin 1 showed elevated expression (p < 0.05). SIRT1 is located in the cell nucleus and plays a vital role in cellular regulation by deacetylating various transcription factors. This activity helps the cell respond to metabolic stressors and modulates cellular longevity and stress resistance, a finding of paramount importance considering the protein’s role in cellular longevity and defence against oxidative stress (7, 13). This upregulation points towards an augmented cellular defence mechanism, potentially leading to enhanced follicle vitality and a prolonged anagen phase. Furthermore, collagen XVIIA expression was significantly increased (p < 0.03) (16). Given that collagen XVIIA is integral for mediating stem cell interactions with the extracellular matrix, this upsurge suggests an improved stem cell niche, fostering enhanced cellular interactions, and potentially promoting hair growth and follicular integrity.
Conversely, a notable reduction was observed in p16INK4A expression (p < 0.03) (14), a biomarker of cellular senescence that underscores a mitigation in the ageing process of the HFs. Lastly, the expression of phosphorylated-S6 was also diminished (p < 0.02) (15). Given that phosphorylated-S6 is an indicator of mTORC1 activity, a main driver of tissue ageing, this highlights a potential dampening in the ageing signalling pathways (Fig. 1).
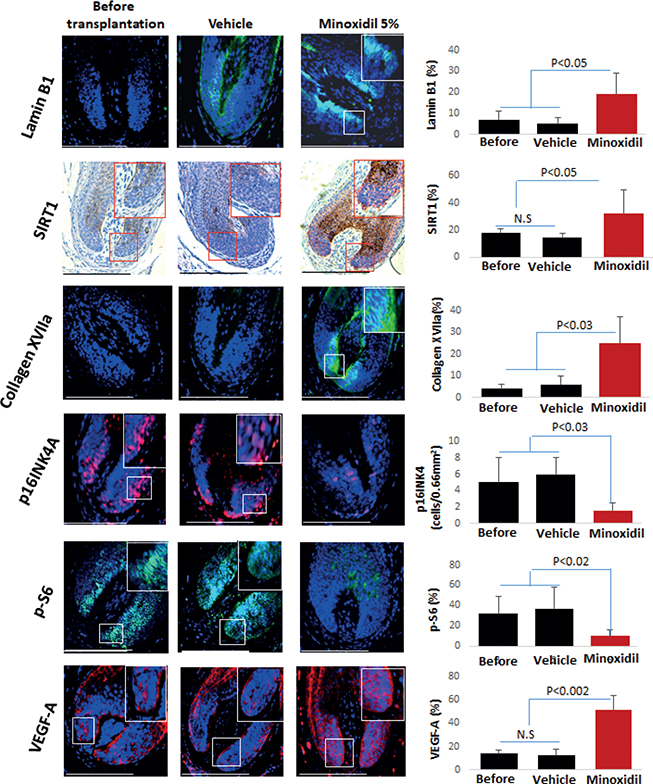
Fig. 1. Minoxidil increased the expression of lamin B1, SIRT1, collagen XVIIA, and VEGF-A and decreased that of protein S6 (p-S6) and p16INK4A in AGA xenotransplants. The graphs show the percentage of positive area in 0.66 mm2 reference areas on 3 sections per sample. SIRT1 (Sirtuin 1) and VEGF-A (vascular endothelial growth factor). Data are mean ± SD. Mann–Whitney U test was applied to compare groups, with p < 0.05 values deemed statistically significant. Scale bar = 50 μm.
Topical minoxidil also significantly enhanced the expression of key proteins related to mitochondrial function in HFs of mpAGA xenotransplants in vivo. Specifically, the treatment notably augmented the expression of peroxisome proliferator-activated receptor gamma coactivator 1-alpha (PGC1α) (p < 0.02), a master regulator of mitochondrial biogenesis and a recognized marker of ageing (17). Reduced PGC1α expression is not merely an indicator of declining mitochondrial function but is also intrinsically linked to the ageing process.
Furthermore, expressions of MTCO1 (p < 0.04) and porin/VDAC (p < 0.05) were also elevated due to the treatment (Fig. S1). These proteins are integral to the optimal functioning of the mitochondria. In our previous study (18), we established a correlation between MTCO1 and porin/VDAC protein expression with the activities of respiratory chain complexes I and IV, underscoring their significance in assessing mitochondrial health and function.
DISCUSSION
These findings underscore the multi-faceted molecular response to minoxidil treatment in mpAGA, highlighted by the upregulation of proteins associated with cellular vitality and the downregulation of ageing markers, illustrating its potential in counteracting ageing effects linked to diminished mitochondrial function.
While the expression of NRF2 itself did not differ significantly between test and control xenotransplants, minoxidil significantly increased the protein expression key NRF2 downstream targets that coordinate oxidative damage responses, namely heme oxygenase-1 (HO-1) (p < 0.01), which is known for its role in the degradation of heme into biliverdin, iron ions, and carbon monoxide, thereby exerting a protective effect against oxidative stress (19). Similarly, peroxiredoxin (PRDX), an enzyme crucial for reducing peroxide levels and mitigating oxidative damage (7), also exhibited enhanced expression (p < 0.03), and glutathione reductase (p < 0.001) along the treated hair bulb (Fig. S2). These antioxidants enhance skin’s resilience against ageing, by bolstering cellular antioxidant defences and reducing oxidative damage.
As VEGF-A is a main driver of human skin rejuvenation in vivo and ex vivo and enhances HO-1 expression (7), we also examined how minoxidil treatment impacted on VEGF-A expression. QIHM revealed that topical minoxidil significantly upregulated VEGF-A protein expression by human outer root sheath keratinocytes in vivo compared with the vehicle-treated group and before the transplantation (p < 0.002) (see Fig. 1). Thus, together with our findings and minoxidil-improved mitochondrial function in vivo (see Fig. S1), and in vitro data that upregulation of VEGF may improve impaired mitochondrial function of mpAGA-derived dermal papilla cells (20), it is conceivable that topical minoxidil can reduce mpAGA-associated intrafollicular oxidative stress and thus slow hair loss progression.
In summary, our data demonstrate that long-term topical application of 5% minoxidil exerts surprisingly widespread rejuvenation effects on aged human HFs within mpAGA scalp skin lesions in vivo on multiple levels. These range from the improvement of core ageing biomarkers (p16INK4, SIRT1, lamin B1, p-S6, collagen XVIIA) and key mitochondrial function read-out parameters (MTCO1, PGC1a, porin/VDAC) to increasing intrafollicular VEGF-A expression and the activation of reparative oxidative damage response pathways (HO-1, PDRX, glutathione transferase) (see Table SII). Subsequent work now needs to dissect mechanistically how minoxidil exerts these unexpected, complex skin and HF rejuvenation effects, given that these can hardly be explained by potassium channel opening alone. Further, it deserves rigorous analysis as to whether low-dose systemic minoxidil, which is increasingly being employed in mpAGA management (21), also exerts at least some anti-ageing effects that mimic those of high-dose topical minoxidil as described here, perhaps even in extracutaneous human organs and tissues. Finally, our data also strongly suggest that topical minoxidil improves the oxidative damage response capacity of human scalp HFs in vivo. This is not only expected to be beneficial in an mpAGA management context (22), but may also award HFs relative protection from oxidative stress, HF stem cell damage, and mitochondrial dysfunction induced by chemotherapy and other drugs (23).
ACKNOWLEDGEMENTS
IRB approval status: IL-150-11-2019.
REFERENCES
- Gupta AK, Carviel J. Androgenetic alopecia: a review. Skin Appendage Disorders 2019; 5: 215–224.
- Chew EGY, Lim TC, Leong MF, Liu X, Sia YY, Leong ST, et al. Observations that suggest a contribution of altered dermal papilla mitochondrial function to androgenetic alopecia. Exp Dermatol 2022; 31: 906–917.
- Matsumura H, Mohri Y, Binh NT, Morinaga H, Fukuda M, Ito M, et al. Hair follicle aging is driven by transepidermal elimination of stem cells via COL17A1 proteolysis. Science 2016; 351: aad4395.
- Haslam IS, Jadkauskaite L, Szabó IL, Staege S, Hesebeck-Brinckmann J, Jenkins G, et al. Oxidative damage control in a human (mini-) organ: Nrf2 activation protects against oxidative stress-induced hair growth inhibition. J Invest Dermatol 2017; 137: 295–304.
- Chew EGY, Tan JHJ, Bahta AW, Ho BS, Liu X, Lim TC, et al. Differential expression between human dermal papilla cells from balding and non-balding scalps reveals new candidate genes for androgenetic alopecia. J Invest Dermatol 2016; 136: 1559–1567.
- Yano K, Brown LF, Detmar M. Control of hair growth and follicle size by VEGF-mediated angiogenesis. J Clin Invest 2001; 107: 409–417.
- Keren A, Bertolini M, Keren Y, Ullmann Y, Paus R, Gilhar A. Human organ rejuvenation by VEGF-A: lessons from the skin. Sci Adv 2022; 8: eabm6756.
- Yum S, Jeong S, Kim D, Lee S, Kim W, Yoo JW, et al. Minoxidil induction of VEGF Is mediated by inhibition of HIF-prolyl hydroxylase. Int J Mol Sci 2017; 19: 53.
- Jadkauskaite L, Coulombe PA, Schäfer M, Dinkova-Kostova AT, Paus R, Haslam IS. Oxidative stress management in the hair follicle: could targeting NRF2 counter age-related hair disorders and beyond? Bioessays 2017; 39.
- Jadkauskaite L, Bahri R, Farjo N, Farjo B, Jenkins G, Bhogal R, et al. Nuclear factor (erythroid-derived 2)-like-2 pathway modulates substance P-induced human mast cell activation and degranulation in the hair follicle. J Allergy Clin Immunol 2018; 142: 1331–1333.
- Gilhar A, Keren A, Ullmann Y, Wu J, Paus R. Effect of minoxidil formulations on human scalp skin xenotransplants on SCID mice: a novel pre-clinical in vivo assay for androgenetic alopecia research. Exp Dermatol 2022; 31: 980–982.
- Laufer Britva R, Keren A, Ginzburg A, Ullmann Y, Paus R, Gilhar A. Evidence from a humanized mouse model of androgenetic alopecia that platelet-rich plasma stimulates hair regrowth, hair shaft diameter and vellus-to-terminal hair reconversion in vivo. Br J Dermatol 2021; 185: 644–646.
- Bulbiankova D, Díaz-Puertas R, Álvarez-Martínez FJ, Herranz-López M, Barrajón-Catalán E, Micol V. Hallmarks and biomarkers of skin senescence: an updated review of skin senotherapeutics. Antioxidants (Basel) 2023; 12: 444.
- Wang AS, Dreesen O. Biomarkers of cellular senescence and skin aging. Front Genet 2018; 9: 247.
- Suzuki T, Chéret J, Scala FD, Akhundlu A, Gherardini J, Demetrius DL, et al. mTORC1 activity negatively regulates human hair follicle growth and pigmentation. EMBO Rep 2023; 24: e56574.
- Matsumura H, Mohri Y, Binh NT, Morinaga H, Fukuda M, Ito M, et al. Hair follicle aging is driven by transepidermal elimination of stem cells via COL17A1 proteolysis. Science 2016; 351: aad4395.
- Abu Shelbayeh O, Arroum T, Morris S, Busch KB. PGC-1α is a master regulator of mitochondrial lifecycle and ROS stress response. Antioxidants (Basel) 2023; 12: 1075.
- Vidali S, Knuever J, Lerchner J, Giesen M, Bíró T, Klinger M, et al. Hypothalamic–pituitary–thyroid axis hormones stimulate mitochondrial function and biogenesis in human hair follicles. J Invest Dermatol 2014; 134: 33–42.
- Ryter SW. Heme oxygenase-1: an anti-inflammatory effector in cardiovascular, lung, and related metabolic disorders. Antioxidants (Basel) 2022; 11: 555.
- Sreedhar A, Aguilera-Aguirre L, Singh KK. Mitochondria in skin health, aging, and disease. Cell Death Dis 2020; 11: 444.
- Randolph M, Tosti A. Oral minoxidil treatment for hair loss: a review of efficacy and safety. J Am Acad Dermatol 2021; 84: 737–746.
- Ntshingila S, Oputu O, Arowolo AT, Khumalo NP. Androgenetic alopecia: an update. JAAD Int 2023; 13: 150–158.
- Piccini I, Brunken L, Chéret J, Ghatak S, Ramot Y, Alam M, et al. Peroxisome proliferator-activated receptor-γ signalling protects hair follicle stem cells from chemotherapy-induced apoptosis and epithelial-mesenchymal transition. Br J Dermatol 2022; 186: 129–141.