SHORT COMMUNICATION
Lesional Infiltration of Eosinophils, Basophils, and M2 Macrophages Expressing Eotaxin-1 and Eotaxin-3 in Granuloma Faciale, but not in Erythema Elevatum Diutinum
Nozomi AKAI#, Takashi HASHIMOTO#*, Manami OKUZAWA, Satoshi OKUNO and Takahiro SATOH
Department of Dermatology, National Defense Medical College, 3-2, Namiki, Tokorozawa, 359-8513 Saitama, Japan. *E-mail: hashderm@ndmc.ac.jp
#These authors contributed equally to this manuscript.
Citation: Acta Derm Venereol 2024; 104: adv35401. DOI: https://doi.org/10.2340/actadv.v104.35401.
Copyright: © Published by Medical Journals Sweden, on behalf of the Society for Publication of Acta Dermato-Venereologica. This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial 4.0 International License (https://creativecommons.org/licenses/by-nc/4.0/)
Submitted: Dec 28, 2023; Accepted: Jan 22, 2024; Published: Feb 12, 2024
Competing interests and funding: The authors have no conflicts of interest to declare.
INTRODUCTION
Erythema elevatum diutinum (EED) and granuloma faciale (GF) are both rare inflammatory skin dermatoses with unknown aetiologies (1, 2). While EED lesions typically develop on the extensor surfaces of the limbs, hands, and fingers, GF principally affects the face. Skin lesions of EED and GF show similar clinical features, characterized by single/multiple persistent erythematous plaques/nodules, although purpuric lesions are more common in EED than GF. Histopathological similarities, such as massive leukocyte infiltration, varying degrees of vascular involvement, and dermal fibrosis, are also found in these diseases (1, 2).
Vascular involvement in EED and GF is usually characterized by leukocytoclastic vasculitis (1, 2). Recent studies have indicated that leukocytoclastic vasculitis involves the formation of extracellular DNA traps (ETs) from neutrophils (3). However, whether or not ETs are formed around vasculature in GF and EED is unclear.
Lesional infiltration of eosinophils intermingled with neutrophils is a crucial histopathological finding that distinguishes GF from EED (1, 2). In general, eosinophils are recruited toward the inflamed sites by C-C chemokines, such as eotaxin-1 (CCL11) and eotaxin-3 (CCL26) (4). Typical cases of EED and GF are experienced at our clinic, and this study aims to evaluate the formation of ETs in vascular lesions. A further aim is to characterize cellular infiltrates expressing etotaxin-1/-3- and/or IL-4/-13.
MATERIALS AND METHODS
Formalin-fixed paraffin-embedded skin samples (5-μm thickness) were deparaffinized and subjected to antigen retrieval using a target retrieval solution (Dako, Glostrup, Denmark) at 60°C overnight. Samples were then treated with Protein Block serum-free (Dako) at room temperature for 10 min, followed by incubation with primary antibodies to citrullinated histone H3 (CitH3) (Catalog #, ab281584, Abcam, Cambridge, UK), eotaxin-1 (ab133604, Abcam), eotaxin-3 (TSC-1; ab217328, Abcam), CD68 (ab955, Abcam), Arg1 (ab92274, Abcam), CD163 (bs-2527R, Bioss antibodies, Woburn, MA, USA), basophil (2D7; ab155577, Abcam), IL-4 (ab9622, Abcam), IL-13 (ab9576, Abcam), and galectin-10 (ab157475, Abcam) at 4°C overnight. Next, the samples were treated with Alexa Fluor 488- or 568-conjugated secondary antibodies and mounted with Fluoroshield with DAPI (GeneTex, Irvine, CA, USA). Samples stained with haematoxylin and eosin (H&E) were also prepared. This study was approved by the institutional review board of the National Defense Medical College (approval number 4477).
RESULTS
Skin samples were collected from 1 patient with EED (a 79-year-old Japanese woman with rheumatoid arthritis) (Fig. 1a) and 1 patient with GF (a 54-year-old otherwise healthy Japanese man) (Fig. 1b). Histopathologically, both EED and GF lesions showed dense cellular infiltrates comprising neutrophils and macrophages with nuclear dust and mild fibrosis in the dermis (Fig. 1c–f). The epidermis was not affected. Eosinophil infiltration was observed only in the GF lesion (Fig. 1f). Due to their massive cell infiltration, it was difficult to identify vascular structures by H&E staining. Thus, immunofluorescence staining was performed. CD31+ endothelial cells were observed lining the dermal vasculature. Notably, these vasculatures were accompanied by formation of neutrophil ETs (NETs), identified with an ET marker CitH3 (5), in both EED and GF lesions (Fig. 1g, h). In addition, eosinophil ETs (EETs) identified by galectin-10 (Fig. 1i) (6) were also detected in GF lesions.
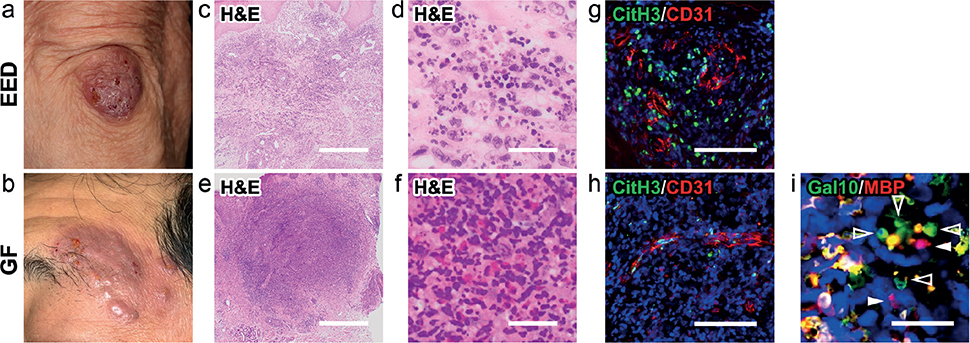
Fig. 1. Clinical features, histopathological findings, and extracellular DNA trap formation in the described cases. (a, c, d, g) Erythema elevatum diutinum (EED). (b, e, f, h, i) Granuloma faciale (GF). (a) Initial presentation of the patient with EED. A brownish firmly elevated nodule was present on the extensor surface of the elbow. (c, d) Haematoxylin and eosin (H&E) staining showed dermal neutrophilic infiltrates with nuclear dust. (b) The GF lesion. Brownish plaques/nodules of various sizes were present on the patient’s left temporal region. (e, f) H&E staining showed massive dermal infiltrates of neutrophils and eosinophils. (g, h) Citrullinated histone H3 (CitH3)-expressing extracellular DNA traps (green) were observed at the vasculatures, as identified by the endothelial marker CD31 (red). (i) ETotic eosinophils with extracellular traps (degenerated DNA, blue), cell-free granules (punctate MBP, red; arrowheads) and extracellular vesicles (punctate galectin-10 [Gal10], green; open arrowheads) were seen. The blue areas indicate nuclei (g, h, i). White bar = 1 mm (c, e), 30 μm (d, f, g, h), and 20 μm (i).
Next, the expression of eotaxin-1 and eotaxin-3 was assessed. In the GF lesion, there was a substantial infiltration of cells expressing eotaxin-1 and eotaxin-3, whereas such infiltration was absent in the EED lesion (Fig. 2a). Moreover, the eotaxin-1- and eotaxin-3-expressing cells were CD68+ macrophages (Fig. 2b). Macrophages are generally divided into 2 subgroups: M1 and M2. M1 macrophages are pro-inflammatory and associated with Th1 immunity, while M2 macrophages are anti-inflammatory and their differentiation is generally promoted by type 2 cytokines, IL-4, and IL-13 (7). Infiltrating CD68+ macrophages in the GF lesion expressed the M2 macrophage markers CD163 and arginase-1 (Fig. 2c) (7, 8). In contrast, the CD68+ cells in the EED lesion did not express M2 macrophage markers (Fig. 2c). Epidermal expression of eotaxin-1 and eotaxin-3 was not observed in both lesions.
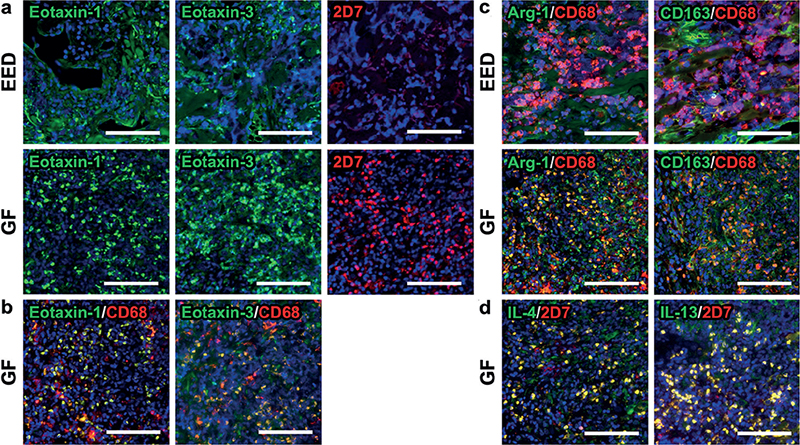
Fig. 2. Immunofluorescence staining findings in the current cases. (a) Cells expressing eotaxin-1 (green in the left panels) and eotaxin-3 (green in the middle panels) were absent in erythema elevatum diutinum (EED), but significantly accumulated in the granuloma faciale (GF) lesion. 2D7+ basophils (red in the right panels) were detected in the GF lesion, but not the EED lesion. (b) Eotaxin-1 (green in the left panel) and eotaxin-3 (green in the right panel) were expressed by CD68+ cells (red) in the GF lesion. (c) CD68+ macrophages (red) expressed M2 macrophage markers Arg-1 (green in the left panels) and CD163 (green in the right panels) in the GF lesion, but not the EED lesion. (d) IL-4 (green in the left panel) and IL-13 (green in the right panel) were mostly expressed by 2D7+ basophils (red) in the GF lesion. The blue areas indicate nuclei. White bar = 100 μm.
Eotaxin-1 and eotaxin-3 function as chemo-attractants not only for eosinophils, but also for basophils, which are known to promote type 2 immunity via secretion of IL-4 and IL-13 (9). In this study, the GF lesion, but not the EED lesion, exhibited a massive infiltration of 2D7+ basophils in the dermis (Fig. 2a). Moreover, 2D7+ basophils expressed IL-4 and IL-13 (Fig. 2d).
DISCUSSION
This study observed the formation of ETs in the vascular lesions of EED and GF. ETs principally originate from neutrophils (NETs) and eosinophils (EETs), and their formation requires histone citrullination. NETs and EETs were initially described as having physiological anti-pathogen functions (5), but are now known to play a role in atherosclerosis, auto-immune disorders (e.g. systemic erythematosus and rheumatoid arthritis), and several types of vasculitis (3, 10, 11). Although the significance of vascular involvement/vasculitis in the pathogenesis of EED and GF remains unclear, controlling NETs and EETs might be a potential therapeutic option for these conditions.
Eotaxin-1 and eotaxin-3 are produced by various types of cells, including epithelial cells, fibroblasts, T lymphocytes, eosinophils, and macrophages (4, 12). In the GF lesion in the current study, eotaxin-1 and eotaxin-3 were found to be expressed by M2 macrophages. A significant number of infiltrating basophils expressed IL-4 and IL-13. Basophils play a pivotal role in recruiting eosinophils to the sites of inflammation in murine contact dermatitis (13) and prurigo-like lesions (14). Basophils in GF lesions might facilitate M2 macrophage-skewing and induce the production of eotaxin-1 and eotaxin-3 by M2 macrophages, which could lead to eosinophil infiltration and further promotion of basophil recruitment. However, the mechanism of the initial basophilic infiltration remains to be elucidated.
Limitations of this study are that only a single case of each disease was examined and only immunofluorescence staining was performed for detecting cellular infiltration and protein expression levels. Nevertheless, the data shown in this study might offer a clue to further understanding the disease entities of GF and EED.
ACKNOWLEDGEMENTS
This study was partially supported by Japan Society for the Promotion of Science (JSPS) KAKENHI Grant-in-Aid for Scientific Research (C) (22K08395 to TH).
This study was approved by the institutional review board of the National Defense Medical College (approval number 4477).
REFERENCES
- Frantz R, Chukwuma O, Sokumbi O, Satcher K, Kallis P, Vincek V, et al. Identifying histopathologic features of erythema elevatum diutinum and granuloma faciale. J Cutan Pathol 2022; 49: 323–326.
- Ziemer M, Koehler MJ, Weyers W. Erythema elevatum diutinum – a chronic leukocytoclastic vasculitis microscopically indistinguishable from granuloma faciale? J Cutan Pathol 2011; 38: 876–883.
- Yousefi S, Simon D, Stojkov D, Karsonova A, Karaulov A, Simon HU. In vivo evidence for extracellular DNA trap formation. Cell Death Dis 2020; 11: 300.
- Amerio P, Frezzolini A, Feliciani C, Verdolini R, Teofoli P, Pità O, et al. Eotaxins and CCR3 receptor in inflammatory and allergic skin diseases: therapeutical implications. Curr Drug Targets Inflamm Allergy 2003; 2: 81–94.
- Hashimoto T, Okuno S, Okuzawa M, Satoh T. Dermal, but not epidermal, immune reaction in cutaneous spiruroid larva migrans: involvement of basophils in dermal interleukin-33 and thymic stromal lymphopoietin expression. Acta Derm Venereol 2022; 102: adv00761.
- Fukuchi M, Kamide Y, Ueki S, Miyabe Y, Konno Y, Oka N, et al. Eosinophil ETosis-mediated release of galectin-10 in eosinophilic granulomatosis with polyangiitis. Arthritis Rheumatol 2021; 73: 1683–1693.
- Yunna C, Mengru H, Lei W, Weidong C. Macrophage M1/M2 polarization. Eur J Pharmacol 2020; 877: 173090.
- Hashimoto T, Satoh T, Yokozeki H. Pruritus in ordinary scabies: IL-31 from macrophages induced by overexpression of thymic stromal lymphopoietin and periostin. Allergy 2019; 74: 1727–1737.
- Karasuyama H, Miyake K, Yoshikawa S, Yamanishi Y. Multifaceted roles of basophils in health and disease. J Allergy Clin Immunol 2018; 142: 370–380.
- Bergqvist C, Safi R, El Hasbani G, Abbas O, Kibbi A, Nassar D. Neutrophil extracellular traps are present in immune-complex-mediated cutaneous small vessel vasculitis and correlate with the production of reactive oxygen species and the severity of vessel damage. Acta Derm Venereol 2020; 100: adv00281.
- Hashimoto T, Ueki S, Kamide Y, Miyabe Y, Fukuchi M, Yokoyama Y, et al. Increased circulating cell-free dna in eosinophilic granulomatosis with polyangiitis: implications for eosinophil extracellular traps and immunothrombosis. Front Immunol 2022; 12: 801897.
- Radonjic-Hoesli S, Brüggen MC, Feldmeyer L, Simon HU, Simon D. Eosinophils in skin diseases. Semin Immunopathol 2021; 43: 393–409.
- Nakashima C, Otsuka A, Kitoh A, Honda T, Egawa G, Nakajima S, et al. Basophils regulate the recruitment of eosinophils in a murine model of irritant contact dermatitis. J Allergy Clin Immunol 2014; 134: 100–107.
- Hashimoto T, Satoh T, Yokozeki H. Protective role of STAT6 in basophil-dependent prurigo-like allergic skin inflammation. J Immunol 2015; 194: 4631–4640.