ORIGINAL REPORT
Inhibitory Immune Checkpoints beyond Programmed Cell Death Ligand 1 in Merkel Cell Carcinoma: Abundant Expression of TIGIT Independent of the Presence of Merkel Cell Polyoma Virus
Ferdinand TOBERER1#, Julia K. WINKLER1#, Leroy ATIENZA FERNANDEZ2,3, Lea ADAMS4§, Alexander BROBEIL5,6, Marcell TÓTH5, Alexander H. ENK1, Jürgen C. BECKER2,3 and Anke S. LONSDORF1
1Department of Dermatology, University Hospital Heidelberg, Heidelberg, 2Departments of Translational Skin Cancer Research and Dermatology, University Hospital Essen, Essen, 3German Cancer Consortium (DKTK), Partner Site Essen/Düsseldorf and German Cancer Research Center (DKFZ), Heidelberg, 4Division of Epigenetics, DKFZ-ZMBH Alliance, German Cancer Research Center, Heidelberg, 5Institute of Pathology, Heidelberg University Hospital, Heidelberg, and 6Tissue Bank of the National Center for Tumor Diseases, Heidelberg, Germany
#These authors contributed equally and share first authorship.
§Current affiliation: Institute of Molecular Health Sciences, Department of Biology, ETH Zurich, Zurich, Switzerland
Merkel cell carcinoma is a rare, aggressive skin cancer in which Merkel cell polyoma virus (MCPyV) is frequently pathogenically involved. After failure of anti-programmed cell death protein 1/programmed cell death ligand 1 immunotherapy, therapeutic options for advanced disease are limited. The contribution of the coinhibitory checkpoint molecule T cell immunoreceptor with immunoglobulin and immunoreceptor tyrosine-based inhibitory motif domain (TIGIT), a regulator of exhausted CD8+ T cells, to the immunosuppressive Merkel cell carcinoma microenvironment is understudied. This study evaluated the immunohistochemical expression of tumour (Tumor Proportion Score, TPS) and infiltrating immune cells (Immune Cell Score, ICS) for programmed cell death ligand 1, TIGIT, its high-affinity receptor CD155, and CD8 in 21 primary Merkel cell carcinoma and 6 metastases. Unlike CD155, TIGIT was abundantly expressed by tumour and immune cells and independent of the MCPyV status, determined by RT-PCR. Programmed cell death ligand 1+ immune cells were significantly increased in TIGIT TPS-positive and MCPyV-positive primary MCC along with significant intercorrelations of programmed cell death ligand 1 and TIGIT immune cell expression and CD8+ infiltrates. Programmed cell death ligand 1 IC-positivity correlated with superior disease-specific survival. The data indicate that TIGIT may contribute to local immune dysfunction in Merkel cell carcinoma, beyond programmed cell death ligand 1 and independent of MCPyV, and provide a rationale to further explore TIGIT as a potential target for Merkel cell carcinoma immunotherapy.
SIGNIFICANCE
Merkel cell carcinoma is a rare, aggressive skin cancer often linked to a virus (Merkel cell polyoma virus). When immunotherapy targeting anti-programmed cell death protein 1/programmed cell death ligand 1 fails, treatment options are limited. This study examined the role of TIGIT, a molecule tied to immune cell exhaustion, and other tumour and immune cell markers in Merkel cell carcinoma samples. Regardless of Merkel cell polyoma virus presence, TIGIT was regularly expressed in Merkel cell carcinoma and metastases. TIGIT immune cell expression correlated with CD8 immune cell density and programmed cell death ligand 1 immune cell expression, a positive prognosticator of survival. The findings propose TIGIT may be a potential target for new combination immunotherapies for Merkel cell carcinoma.
Key words: immune checkpoint inhibition; Merkel cell carcinoma; PD-L1; TIGIT; CD155; T cell exhaustion.
Citation: Acta Derm Venereol 2025; 105: adv42882. DOI: https://doi.org/10.2340/actadv.v105.42882.
Copyright: © 2025 The Author(s). Published by MJS Publishing, on behalf of the Society for Publication of Acta Dermato-Venereologica. This is an Open Access article distributed under the terms of the Creative Commons Attribution-NonCommercial 4.0 International License (https://creativecommons.org/licenses/by-nc/4.0/).
Submitted: Jan 14, 2025. Accepted after revision: Jun 2, 2025. Published: Jul 1, 2025.
Corr: Anke S. Lonsdorf, Department of Dermatology, University Hospital Heidelberg, Im Neuenheimer Feld 440, DE-69120 Heidelberg, Germany. E-mail: anke.lonsdorf@med.uni-heidelberg.de
Competing interests and funding: JKW received travel expenses and/or honoraria from Almirall, Amgen, Biofrontera, Biotest, BMS, Fotofinder, Leo Pharma, MSD, Philochem, and Roche. AEH receives funding through grant SFB/TR 156 of the DFG (Deutsche Forschungsgemeinschaft) and payment or honoraria from Janssen-Cilag, Biotest, MSD, and Galderma. AEH is president of the European Dermatology Forum (EDF). JCB is receiving speaker’s bureau honoraria from Amgen, MerckSerono, Pfizer, Sanofi, and Sun Pharma and is a paid consultant/advisory board member/DSMB member for Almirall, Boehringer Ingelheim, ICON, Pfizer, and Sanofi/Regeneron. His group receives research grants from Merck Serono/IQVIA, Regeneron and Alcedis. ASL received travel expenses and honoraria from the Professor Paul Gerson Unna Akademie e.V.. All other authors declare that there is no conflict of interest.
INTRODUCTION
Merkel cell carcinoma (MCC) is a rare, highly aggressive epithelial skin cancer with differentiation that characteristically occurs in chronically ultraviolet (UV)-exposed body sites of elderly and/or immunosuppressed patients (1–3). Apart from UV radiation, Merkel cell polyoma virus (MCPyV) is a chief pathogenic driver in approximately 70–80% of MCC cases (2–4).
Considered an immunogenic cancer, MCC immunogenicity is thought to be largely conveyed by the constitutive expression of oncogenic MCPyV-encoded proteins, i.e., small and large transforming antigen, in McPyV-positive (VP)-MCC and a high mutational burden due to UV exposure in MCPyV-negative (VN)-MCC (1, 2, 5). Brisk immune cell infiltration, particularly with CD8+ tumour infiltrating lymphocytes (TIL), is observed in some MCC and has been reported to favourably influence clinical outcomes in some studies (2, 5–9). However, various inhibitory immune checkpoints (iICP) expressed by solid cancers and their tumour microenvironment (TME) may contribute to local immune dysfunction by inducing exhaustion of CD8+ cytotoxic T cells and other anti-tumour effector immune cells (10–14).
Until recently effective therapeutic options for advanced-stage MCC have been limited, resulting in a poor prognosis and high disease-specific mortality (2, 3, 15). The identification of iICP and their targeted interruption by therapeutic antibodies led to the establishment of effective immunotherapies for multiple advanced cancers, including MCC. Monotherapy targeting the iICP PD-L1/PD-1 evolved as standard of care for advanced-stage MCC, demonstrating effective and durable anti-tumour responses in both VP-MCC and VN-MCC (1, 16, 17).
However, while treatment outcomes of immune checkpoint inhibitors (ICIs) are impressive, primary and secondary resistance remains a challenge in almost two-thirds of advanced MCC patients failing to respond to therapeutic PD-L1/PD-1 blockade (1–3, 15–17). Further, unlike in other cancers, quantification of PD-L1 expression by tumour cells and/or immune cells does not predict response to ICIs in MCC (9, 15), indicating the need for identification of predictive biomarkers and novel targets for conceivably combined therapeutic strategies for advanced-stage MCC (3, 18).
A central role in impeding anti-tumour responses has recently been assigned to a T cell immunoreceptor with immunoglobulin and immunoreceptor tyrosine-based inhibitory motif (ITIM) domain (TIGIT). TIGIT is a member of the immunoglobulin superfamily with well-documented expression on T cells and NK cells (14, 19, 20). In addition to its roles in other immune cells, TIGIT is considered a marker of exhausted CD8+ T cells and may negatively regulate effector immune responses by several mechanisms, including directly transmitting cell-intrinsic inhibitory signals or by interacting with its high-affinity ligand CD155 (poliovirus receptor, PVR) (14, 19, 20).
The TIGIT/CD155 axis has been tied to resistance to ICIs in melanoma (21) and we and others recently highlighted TIGIT/CD155 as potential contributors to immune exhaustion and as possible new therapeutic targets in different cancers (14, 22–28). Yet, expression of TIGIT/CD155 by rare skin cancers, such as MCC, is not well established with only 1 single study examining the prognostic value of Forkhead helix transcription factor P3 (FOXP3) and the TIGIT/CD155 axis in MCC of Asian patients (6).
In this study, we comprehensively assessed the collective landscape of immunomorphology patterns of PD-L1, TIGIT, and CD155 of both tumour and immune cells in primary MCC and cutaneous metastases. We further disclose reciprocal correlations of these iICP and examined their potential association with clinicopathological parameters, including the presence of MCPyV and disease-specific survival. Our study provides a rationale for further investigating the potential of ICI targeting TIGIT in MCC.
MATERIALS AND METHODS
Patients and samples
A total of 27 tissue samples from 26 patients were analysed, comprising 21 primary MCC and 6 cutaneous metastases. All patients underwent routine surgery for their tumours or metastases at the Department of Dermatology, University Hospital of Heidelberg, Germany. Included patients were not treated with ICIs prior to surgery. Clinicopathological parameters, including age at diagnosis, sex, tumour site, stage, and survival were determined through medical records and preoperative photographs. Diagnoses and histopathologic features of all samples were determined by board-certified dermatopathologists. Formalin-fixed paraffin-embedded (FFPE) samples were retrieved from the tissue bank of the Department of Dermatology, University Hospital Heidelberg (Biobank of the National Center for Tumor Diseases Heidelberg, Germany) in accordance with institutional guidelines. The study was approved by the ethics committee of Heidelberg University and all participants gave prior written informed consent.
Immunohistochemistry
Immunohistochemistry (IHC) of serial 3μm FFPE tissue sections was performed at the Biobank of the NCT Heidelberg, as previously described (23), on an automated immunostainer (Ventana BenchMark Ultra, Ventana Medical Systems, Oro Valley, AZ, USA) with monoclonal antibodies against PD-L1 (clone SP263, Roche, Basel, Switzerland) and CD8 (clone SP57, Roche) at the provided dilutions of the ready-to-use kits and a monoclonal anti-CD155 antibody (clone D8A5G, Cell Signaling Technology, Danvers, MA, USA) at 1:200 dilution and visualization by the biotin-free OptiView DAB IHC Detection Kit (Ventana Medical Systems). TIGIT IHC was performed after antigen retrieval in citrate buffer (pH 6.0) with a monoclonal anti-TIGIT antibody (clone TG1, OncoDianova, Hamburg, Germany) at 1:100 dilution on a Bond RX automated immunostainer (Leica Biosystems, Nussloch, Germany) with AEC-Chromogen (Dako REAL EnVision Detection System, Dako, Agilent Technologies Inc, Santa Clara, CA, USA). Germinal centres of normal human tonsil tissue, showing high TIGIT expression (29), served as positive control for TIGIT IHC, while colon carcinoma served as a positive control for CD155 expression (28). Slides were counterstained with hemalaun.
Evaluation of immunostaining
In view of reported considerable intratumoral heterogeneity for iICP expression (30), entire tissue slides were analysed. Evaluation of immunostaining was performed as previously described (23) by a board-certified dermatopathologist blinded to clinical parameters. Staining of the tumour cell component and immune cells in the intra- and peritumoral stroma were analysed separately and expressed as Tumor Proportion Score (TPS), defined as the percentage of viable tumour cells exhibiting complete or partial membranous staining at any intensity above background cytoplasmic staining (PD-L1 and CD 155) or percentage of viable tumour cells exhibiting cytoplasmic staining at any intensity (TIGIT), and as Immune Cell Score (ICS), as previously defined for PD-L1, respectively (31). For TIL, both cytoplasmatic and membranous staining were considered for positivity. Based on definitions used in the registrational trials for ICI in MCC, PD-L1 TPS positivity was defined as ≥ 1% stained tumour cells (16–18) and PD-L1 IC positivity as ≥ 1% stained infiltrating immune cells. CD8 immunostainings were regarded as positive in the case of specific cytoplasmic or membranous staining and expressed as the proportion of positive cells relative to the tumour cell area. The median value was used as cutoff to distinguish between high and low densities of CD8+ immune infiltrates. CD155 immunostaining was considered positive only in the case of a specific membranous staining and expressed as percentage of analysed cells, as previously described (22). Cut-off values for TIGIT and CD155 positivity in MCC are not clearly established and were therefore set at ≥ 5% and ≥ 1%, respectively, for statistical analysis, as previously suggested (2, 23, 27).
MCPyV status
MCPyV status was determined by TaqMan Real-Time qPCR as previously described, using the following large T-antigen specific primers and TaqMan probe: forward primer, CCA_ AAC_CAA_AGA_ATA_AAG_CAC_TGA; reverse primer, TCG_CCA_GCA_TTG_TAG_TCT_AAA_AAC, and probe: FAM-AGC_AAA_AAC_ACT_CTC_CCC_ACG_TCA_GAC_AG-BHQ1 (32).
Statistical analyses
Analysis of immunoexpression between dichotomous groups defined by MCPyV status and immunoexpression status was performed by exact Mann–Whitney U test using R version 4.3.0 (2023-04-21; package “coin” version 1.4-3; R Foundation for Statistical Computing, Vienna, Austria) (33). IBM SPSS Statistics (version 29.0.0.0; IBM Corp, Armonk, NY, USA) was used for all other analyses. Clinicopathological characteristics between dichotomous groups defined by immunoexpression status were analysed by Fisher’s exact test or Mann–Whitney U test (for age). Differences in the quantitative expression between MCC and metastases were assessed by Mann–Whitney U test. Spearman’s rank correlation was used to analyse the association of quantitative expression measures. As analyses were exploratory, no adjustments for multiple comparisons were performed. Disease-specific survival (DSS) was analysed by Kaplan–Meier estimator and log-rank test to compare distributions between groups. P < 0.05 was considered indicative of statistical significance.
RESULTS
Clinicopathological characteristics
Baseline clinicopathological characteristics are presented in Table I. The majority of patients were male (53.8%). The median age at diagnosis was 80.7 years (range: 52.1–92.1) and 50% of patients were initially diagnosed with stage I MCC. Three of the 6 analysed metastases, but no primary tumour, originated from immunosuppressed kidney transplant recipients. Tumours were typically located in chronic UV-exposed sites on the upper extremities (40.7%) or the head/neck area (29.6%) and were mostly classified as VP-MCC (55.6%).
| Sex, years, n (%) | |
| Male | 14 (53.8) |
| Female | 12 (46.2) |
| Age, years, median (range) | 80.7 (52.1–92.1) |
| Tissue, n (%) | |
| MCC | 21 (77.8) |
| MCC metastasis | 6 (22.2) |
| Anatomic site, n (%) | |
| Head/neck | 8 (29.6) |
| Upper extremities | 11 (40.7) |
| Lower extremities | 4 (14.8) |
| Trunk | 4 (14.8) |
| Immunosuppression, n (%) | |
| Yes /No | 3 (11.5) / 23 (88.5) |
| AJCC stagea, n (%) | |
| I/II | 13 (50.0) / 8 (30.8) |
| III/IV | 5 (19.2) / 0 (0) |
| Disease-specific survival, months, mean (95% CI) | 31.3 (25.1–37.6) |
| MCPyV status, n (%) | |
| Negative | 12 (44.4) |
| Positive | 15 (55.6) |
| aAt the time of primary diagnosis, according to the American Joint Committee on Cancer staging (AJCC) for Merkel cell carcinoma, 8th edition. Clinical and pathological stages combined. Mean disease-specific survival (DSS) representing the time from histological diagnosis to MCC-specific death with 95% confidence intervals (CI). MCC: Merkel cell carcinoma, MCPyV: Merkel cell polyoma virus. | |
Immunohistochemical iICP expression and CD8+ infiltrates in MCC and metastases
Representative IHC results are shown in Fig. 1. PD-L1 immunoexpression by infiltrating mononuclear immune cells was detected in the majority of samples (22/27, PD-L1 IC: median: 10%, range: 0–50%), while only 1 primary MCC displayed specific tumoral PD-L1 expression (PD-L1 TPS:1%). TIGIT was abundantly expressed by mononuclear immune cells (TIGIT IC: median: 5%, range: 0–30%) and by tumour cells with cytoplasmic immunostaining (TIGIT TPS: median: 20%, range: 0–100%). By contrast, specific membranous CD155 staining of tumour cells was observed in only 1 MCC (CD155 TPS: 50%). CD8+ immune cell infiltrates were detected in all samples with varying densities (median: 10%, range: 1–70%).

Fig. 1. Immunohistochemical staining. Representative examples of primary Merkel cell carcinoma with immunohistochemical expression of programmed death-ligand 1 (PD-L1) of immune cell infiltrates. (A) PD-L1 Immune Cell Score (ICS) of whole tumour section: 30%), tumour cell expression of T-cell immunoreceptor with immunoglobulin and immunoreceptor tyrosine-based inhibition motif domain (TIGIT). (B) left panel: tumoral TIGIT expression of whole tumour section, 60%, immune cell expression of TIGIT; right panel: TIGIT IC of whole tumour section, 10%, CD155 immunoexpression of tumour cells. (C) CD155 expression of whole tumour section, 50% and CD8+ infiltrating immune cells. (D) CD8+ tumour infiltrating lymphocytes (TIL) density of whole tumour section, 20%. Selected tissue slides were digitalized using an Aperio AT2 Slide scanner (Leica Biosystems, Nussloch, Germany) for image acquisition with Aperio Image Scope Software (version 12.4.0.5043, Leica Biosystems). Original magnification ×100 (upper panels; scale bars: 300 μm) and insert ×400 (lower panels; scale bars: 60 μm).
While no significant differences in iICP immunoexpression between primary MCC and metastases were noted, there was a trend for primary MCC to exhibit overall denser infiltration with CD8+ (p = 0.079) and TIGIT+ (p = 0.091) immune cells (Fig. 2).
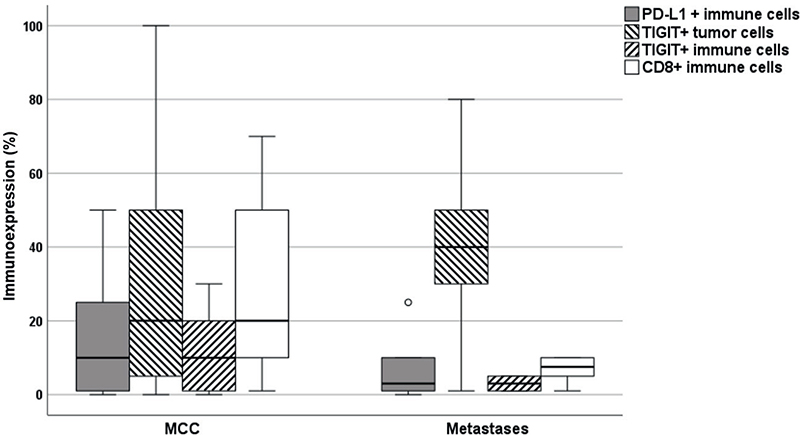
Fig. 2. Immunoexpression of PD-L1 and TIGIT and density of CD8+ immune infiltrates in primary Merkel cell carcinoma and cutaneous metastases. P-values are obtained by Mann–Whitney U test. P < 0.05 was considered statistically significant. MCC: Merkel cell carcinoma, PD-L1: programmed death-ligand 1, TIGIT: T cell immunoreceptor with immunoglobulin and immunoreceptor tyrosine-based inhibitory motif.
Associations of iICP expression status and CD8+ profiles with clinicopathological parameters
First, we analysed whether iICP positivity at indicated cut-offs or the density of CD8+ immune cell infiltrates were associated with clinicopathological features (Table II). While only 1 primary MCC displayed specific tumoral PD-L1 at a ≥ 1% cut-off, the vast majority of samples exhibited PD-L1 IC positivity (81.5%). TIGIT TPS positivity (77.8%) and TIGIT IC positivity (66.7%) was detected in the majority of samples, particularly in primary MCC (TIGIT TPS positivity: 16/21; TIGIT IC positivity: 15/21). Some 50% of MCC metastasis (3/6) and the majority of primary MCC (16/21, 76.2%) displayed high CD8+ immune cell infiltration.
Sex and age were neither associated with iICP immunoexpression nor CD8+ TIL density. No association with clinicopathological parameters was noted for TIGIT positivity, CD155 positivity or CD8+ TIL density. PD-L1 IC positivity was significantly associated with the presence of MCPyV with all VP-MCC exhibiting a positive PD-L1 IC status (p = 0.01). Further, most samples with positivity for PD-L1 IC (15/22, 68.2%), TIGIT IC (11/18, 61.1%), and TIGIT TPS (13/21, 61.9%), as well as the only CD155 expressing MCC, were MCPyV positive.
Associations of MCPyV-status, iICP expression, and CD8+ immune cell density
Taking into account the high frequency of VP-MCC with a positive iICP status at the indicated cut-offs (see Table II) and the reported impact of viral antigens on iICP expression by tumour or immune cells (34, 35), we further stratified samples according to MCPyV status to examine differences in iICP expression and CD8+ immune cell density between VN- and VP-MCC (Fig. 3). In primary MCC, there was no significant difference regarding tumoral and immune cell expression of TIGIT in VP-MCC and VN-MCC (Fig. 3B and C). However, in VP-MCC, PD-L1+ (Fig. 3A, p = 0.003) and CD8+ (Fig. 3D, p = 0.043) immune cell infiltrates were significantly enhanced compared with VN-MCC. In MCC metastases no significant association of iICP expression or CD8+ infiltrates with MCPvV status were found (Fig. S1).
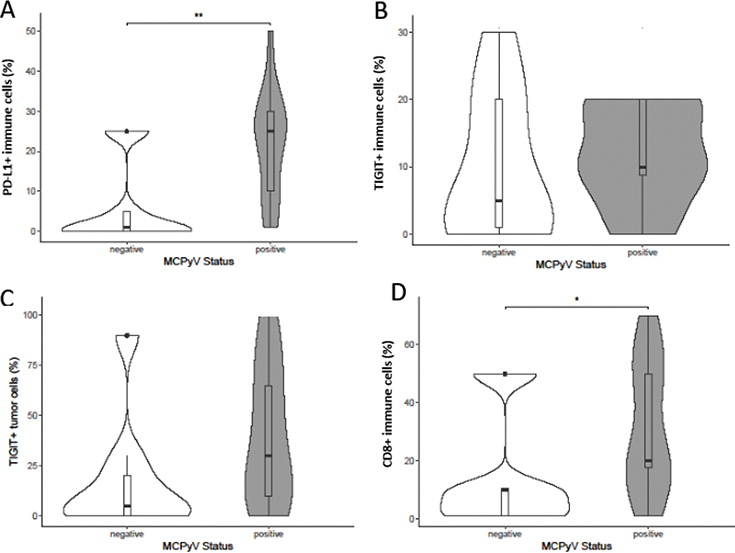
Fig. 3. (A–D) Inhibitory immune checkpoint expression and CD8+ host immunity stratified for the presence of Merkel cell polyoma virus (MCPyV) in primary Merkel cell carcinoma. P-values are obtained by exact Mann–Whitney U test; p < 0.05 was considered statistically significant; *p < 0.05, **p < 0.01. PD-L1: programmed death-ligand 1, TIGIT: T cell immunoreceptor with immunoglobulin and immunoreceptor tyrosine-based inhibitory motif.
Reciprocal relationships between iICP status and CD8+ host immunity
Samples were stratified for their iICP status and CD8+ TIL density at indicated cut-offs to examine their reciprocal relationships. In PD-L1 IC-positive tumours, CD8+ (Fig. 4A, p = 0.001) and TIGIT+ (Fig. 4B, p = 0.002) immune cell infiltrates were significantly increased. TIGIT IC-positive MCC displayed significantly increased CD8+ (Fig. 4C, p = 0.0002) and PD-L1+ (Fig. 4D, p = 0.005) immune cell infiltrates compared with TIGIT IC-negative primary tumours. MCC with high CD8+ density displayed significantly higher immune cell expression of PD-L1 (Fig. 4E, p = 0.0005) and TIGIT (Fig. 4F, p = 0.0001) compared with MCC with low CD8+ immune cell density. While stratification for TIGIT TPS positivity neither exhibited significant differences regarding the density of CD8+ (Fig. 4G) nor TIGIT+ immune cell infiltrates (Fig. 4I), PD-L1+ immune cell infiltrates were significantly enriched in TIGIT TPS-positive MCC (Fig. 4H, p = 0.009).

Fig. 4. Reciprocal relationships between inhibitory immune checkpoint expression patterns and CD8+ host immunity in primary Merkel cell carcinoma. (A) PD-L1 immune cell expression status (IC)/CD8 immunoexpression (%). (B) PD-L1 IC/TIGIT immune cell immunoexpression (%). (C) TIGIT IC/CD8+ immune cells (%). (D): TIGIT IC/PD-L1+ immune cells (%). (E) CD8+ tumour infiltrating lymphocytes (TIL) density/PD-L1+ immune cells (%). (F) CD8+ TIL density/TIGIT+ immune cells (%). (G) TIGIT tumour cell expression status (TPS)/CD8 immunoexpression (%). (H) TIGIT TPS/PD-L1+ immune cells (%). (I) TIGIT TPS/TIGIT immune cell immunoexpression (%); cut-off values: PD-L1 IC: negative: < 1%, positive ≥ 1%; TIGIT IC: negative: < 5%, positive ≥ 5%; TIGIT TPS: negative: < 5%, positive ≥ 5%; CD8+ low: < median, CD8+ high: ≥ median. P-values are obtained by exact Mann–Whitney U test. P < 0.05 was considered statistically significant; **p < 0.01, ***p < 0.001. IC: Immune Cell Score, PD-L1: programmed death-ligand 1, TIGIT: T cell immunoreceptor with immunoglobulin and immunoreceptor tyrosine-based inhibitory motif, TPS: Tumor Proportion Score.
Correlation of iICP expression and CD8+ infiltrates
To gain additional insights into iICP expression patterns we examined the correlations of dichotomous iICP expression and CD8+ TIL densities by Spearman’s rank correlation. PD-L1 immune cell expression positively correlated with the density of CD8+ immune cells (Fig. 5A, p = 0.0002, r = 0.731) and TIGIT+ immune cells (Fig. 5B, p = 0.0006, r = 0.684) in MCC. Further, a strong positive correlation of TIGIT immune cell expression with CD8+ infiltrates was detected (Fig. 5C, p = 0.00001, r = 0.832). Yet, no significant correlations were noted between TIGIT+ tumour cells and PD-L1+, TIGIT+, or CD8+ immune cell infiltrates (Fig. 5D, E and F). Overall no significant correlations between iICP expression patterns and CD8+ infiltrates were noted in MCC metastases (data not shown).

Fig. 5. (A–F) Correlation of iICP co-expression patterns and CD8+ infiltrates in primary Merkel cell carcinoma. Spearman’s rank correlation was used. P < 0.05 was considered statistically significant (in bold), iICP: inhibitory immune checkpoint, PD-L1: programmed death-ligand 1, TIGIT: T cell immunoreceptor with immunoglobulin and immunoreceptor tyrosine-based inhibitory motif.
Disease-specific survival (DSS) stratified by clinicopathological parameters
Mean DSS among 21 immunocompetent patients with primary MCC was 31.3 months (95% CI 25.1–37.6). While sex was not found to be a prognosticator (p = 0.592) (Fig. 6A), DSS was significantly associated with tumour stage (p < 0.014, Fig. 6B). A positive PD-L1 IC status was significantly associated with longer DSS (7.9 months, 95% CI 4.6–11.1 vs 33.2 months, 95% CI 27.6–38.7) (p = 0.011, Fig. 6D). No significant associations of DSS with MCPyV-status (p = 0.175, Fig. 6C), TIGIT TPS (p = 0.361, Fig. 6E), TIGIT IC (p = 0.741, Fig. 6F), or the density of CD8+ immune cells (p = 0.361, Fig. 6G) were found.

Fig. 6. Kaplan–Meier analysis of disease-specific survival (DSS) of patients with primary Merkel cell carcinoma. DSS stratified by (A) sex; (B) tumour stage (American Joint Committee on Cancer staging for Merkel Cell Carcinoma, 8th edition, clinical and pathological stages combined); (C) Merkel cell polyoma virus (MCPyV) status; (D) PD-L1 immune cell expression status (IC); (E) TIGIT tumour cell expression status (TPS); (F) TIGIT immune cell expression status (IC); and (G) density of CD8+ immune cells. PD-L1: programmed death-ligand 1, TIGIT: T cell immunoreceptor with immunoglobulin and immunoreceptor tyrosine-based inhibitory motif. Log-rank test was used to evaluate statistical significance. P < 0.05 was considered statistically significant (in bold).
DISCUSSION
Solid cancers, including MCC, exhibit highly suppressive microenvironments in which iICP such as PD-1/PD-L1 mediate T cell exhaustion and local immune dysfunction (10, 11, 36). Although anti-PD-1/PD-L1 therapy greatly improved outcomes of patients with advanced MCC, various mechanisms may contribute to resistance to ICI, such as loss of MHC class I or neoantigen expression and upregulation of coinhibitory molecules like TIGIT/CD155 (10, 36, 37).
In this study, we detected abundant tumoral TIGIT immunoexpression in a substantial proportion of primary MCC and metastases, independent of MCPyV. Contrasting the well-documented TIGIT expression on immune cells (14, 19, 36) and its central role in mediating T cell exhaustion, the significance of TIGIT expression by tumour cells is currently less clear. Few studies have previously reported TIGIT expression by human cancer cells, specifically melanoma, undifferentiated pleomorphic sarcoma, and most recently MCC (6, 21, 38).
While a recent study by Iwasaki et al. (6) focused on TIGIT expression by MCC tumour cells, we also comprehensively evaluated whole-tissue sections for the presence of TIGIT+ immune cells, revealing abundant TIGIT+ infiltrating mononuclear immune cells in a large proportion of MCC. Consistent with reported associations of TIGIT+ CD8+ infiltrates with increased expression of other iICP (22, 23, 35), our analyses revealed significant enrichment of CD8+ and PD-L1+ immune cells in TIGIT IC-positive tumours. Moreover, significantly increased expression of both PD-L1 and TIGIT on immune cells was detected in MCC with brisk CD8+ immune cell infiltrate along with a consistently observed positive reciprocal correlation of these markers on immune cells. Confirming and complementing the study by Iwasaki et al. (6), reporting overall weak immunostaining of MCC tumour cells for the high-affinity TIGIT receptor CD155, but significantly higher CD155 mRNA expression by VP-MCC, in our study the only MCC with substantial CD155 expression was classified MCPyV-positive.
DSS was not associated with MCPyV in our study and current therapeutic strategies for advanced-stage MCC, including ICI, are currently unaffected by the MCPyV status (3, 18, 39). Nevertheless, iICP expression on tumour or immune cells and TIL density may be affected by the presence of viral antigens in solid cancers (12, 34) and MCPyV-specific CD8+ T cells are frequently detected in patients with VP-MCC (5, 11, 40).
Our study did not encompass functional analyses and further investigation on the abundance and avidity of MCPyV-specific T cells in the TME of VP-MCC is needed (3). Yet, consistent with previous studies (6, 8, 12), we detected a significant enrichment of CD8+ and PD-L1+ immune cells in VP-MCC, possibly indicating that distinct iICP-mediated immune cell exhaustion may occur in response to persistent virus infection. Correspondingly, the only primary MCC displaying specific tumoral PD-L1 or CD155 immunoexpression, respectively, were both classified as VP-MCC. By contrast, we found TIGIT to be expressed in both VN-MCC and VP-MCC on tumour and immune cells. While tumoral TIGIT expression was not associated with increased CD8+ immune cell density or TIGIT+ immune cells, we detected significantly increased PD-L1+ immune cell infiltrates in TIGIT TPS-positive MCC compared with TIGIT TPS-negative MCC. Highlighting TIGIT as a particularly interesting therapeutic target of oncovirus-associated cancers, such as VP-MCC, Johnston et al. demonstrated that TIGIT expression by immune cells collaborates with PD-1/PD-L1 to potently limit the effector function of chronically stimulated CD8+ T cells in both cancer and chronic viral infection (35).
TIGIT inhibitors, such as tiragolumab, vibostolimab, domvanalimab, and ociperlimab, are expected to be most efficient in co-blockade with other iICPs. In particular, combination with anti-PD-1/PD-L1 agents demonstrated superior response rates in preclinical and clinical studies and bispecific antibodies are currently in clinical development (20, 24, 26, 27, 36, 37). The intercorrelations between PD-L1, TIGIT, and CD8+ infiltrates seen in our study are in line with the current perception that tumours and their TME regularly co-express multiple iICPs for immune evasion (11, 20, 22, 23, 36) and indicate that iICP patterns of tumours and their TME may differentially be affected by the presence of viral oncogenes.
Limitations
Limitations of our study merit note, including potential racial and regional variances, as analysis was restricted to a German patient cohort at a single institution. Given the small sample size statistical analysis is largely descriptive and adjustments for multiple comparisons were not performed. Particularly for analyses of survival and metastases, interpretation of results should consider the possibility that subtle but biologically relevant changes may not have been detected. Sample size further hampered examination of an effect of iatrogenic immunosuppression, a relevant clinical prognosticator present in 3/6 investigated metastases. Further restrictions in the robustness of drawn conclusions arise from the use of cut-off values for immunoexpression without complementary continuous validation. Larger-scale studies are needed to validate our findings, to establish cut-offs for TIGIT/CD155 immunoexpression and decisively examine their prognostic relevance in MCC.
Conclusion
Complementing the current understanding on the expression of the iICP TIGIT/CD155 in cutaneous malignancies, our study reveals frequent expression of TIGIT by tumour and immune cells in in both VN-MCC and VP-MCC and deciphers significant intercorrelations of PD-L1 and TIGIT immunoexpression and CD8+ infiltrating immune cells. The biological and clinical significance of TIGIT expression by both tumour and immune cells in the absence of robust expression of its high-affinity receptor CD155, as confirmed for MCC in our study, is currently incompletely understood. Yet, the combined findings of previously reported data (6) and our study together with promising efficacy of a TIGIT/PD-L1 combination therapy in other cancers (20, 24, 26, 27) provide a rational to further investigate TIGIT as a therapeutic target in both VN- and VP-MCC.
ACKNOWLEDGEMENTS
The authors thank the participating patients and staff of the Department of Dermatology at the University Hospital Heidelberg for their valuable contribution to this research and Fabio Tabone and Veronika Eckel, Biobank of the National Center for Tumor Diseases Heidelberg, and Frauke Furtmann, for expert technical assistance.
IRB approval status: Reviewed and approved by the Ethics Committee of the Medical Faculty of the University of Heidelberg; approval Nr. S-091/2011.
REFERENCES
- Gauci ML, Aristei C, Becker JC, Blom A, Bataille V, Dreno B, et al. Diagnosis and treatment of Merkel cell carcinoma: European consensus-based interdisciplinary guideline – Update 2022. Eur J Cancer 2022; 171: 203–231. https://doi.org/10.1016/j.ejca.2022.03.043
- Becker JC, Stang A, DeCaprio JA, Cerroni L, Lebbe C, Veness M, et al. Merkel cell carcinoma. Nat Rev Dis Primers 2017; 3: 17077. https://doi.org/10.1038/nrdp.2017.77
- Harms PW, Harms KL, Moore PS, DeCaprio JA, Nghiem P, Wong MKK, et al. The biology and treatment of Merkel cell carcinoma: current understanding and research priorities. Nat Rev Clin Oncol 2018; 15: 763–776. https://doi.org/10.1038/s41571-018-0103-2
- Feng H, Shuda M, Chang Y, Moore PS. Clonal integration of a polyomavirus in human Merkel cell carcinoma. Science 2008; 319: 1096–1100. https://doi.org/10.1126/science.1152586
- Ouyang K, Zheng DX, Agak GW. T-cell mediated immunity in merkel cell carcinoma. Cancers (Basel) 2022; 14: 6058. https://doi.org/10.3390/cancers14246058
- Iwasaki T, Hayashi K, Matsushita M, Nonaka D, Matsumoto T, Taniguchi M, et al. Clinical significance of the expression of FOXP3 and TIGIT in Merkel cell carcinoma. Sci Rep 2023; 13: 13114. https://doi.org/10.1038/s41598-023-40050-7
- Naseri S, Steiniche T, Georgsen JB, Thomsen R, Ladekarl M, Heje M, et al. Tumor ulceration, reduced infiltration of CD8-lymphocytes, high neutrophil-to-CD8-lymphocyte ratio and absence of MC virus are negative prognostic markers for patients with merkel cell carcinoma. Cancers (Basel) 2020; 12: 888. https://doi.org/10.3390/cancers12040888
- Feldmeyer L, Hudgens CW, Ray-Lyons G, Nagarajan P, Aung PP, Curry JL, et al. Density, distribution, and composition of immune infiltrates correlate with survival in merkel cell carcinoma. Clin Cancer Res 2016; 22: 5553–5563. https://doi.org/10.1158/1078-0432.CCR-16-0392
- Spassova I, Ugurel S, Kubat L, Zimmer L, Terheyden P, Mohr A, et al. Clinical and molecular characteristics associated with response to therapeutic PD-1/PD-L1 inhibition in advanced Merkel cell carcinoma. J Immunother Cancer 2022; 10: e003198. https://doi.org/10.1136/jitc-2021-003198
- Andrews LP, Yano H, Vignali DAA. Inhibitory receptors and ligands beyond PD-1, PD-L1 and CTLA-4: breakthroughs or backups. Nat Immunol 2019; 20: 1425–1434. https://doi.org/10.1038/s41590-019-0512-0
- Schadendorf D, Nghiem P, Bhatia S, Hauschild A, Saiag P, Mahnke L, et al. Immune evasion mechanisms and immune checkpoint inhibition in advanced Merkel cell carcinoma. Oncoimmunology 2017; 6: e1338237. https://doi.org/10.1080/2162402X.2017.1338237
- Lipson EJ, Vincent JG, Loyo M, Kagohara LT, Luber BS, Wang H, et al. PD-L1 expression in the Merkel cell carcinoma microenvironment: association with inflammation, Merkel cell polyomavirus and overall survival. Cancer Immunol Res 2013; 1: 54–63. https://doi.org/10.1158/2326-6066.CIR-13-0034
- Walsh NM, Fleming KE, Hanly JG, Dakin Hache K, Doucette S, Ferrara G, et al. A morphological and immunophenotypic map of the immune response in Merkel cell carcinoma. Hum Pathol 2016; 52: 190–196. https://doi.org/10.1016/j.humpath.2016.02.002
- Manieri NA, Chiang EY, Grogan JL. TIGIT: a key inhibitor of the cancer immunity cycle. Trends Immunol 2017; 38: 20–28. https://doi.org/10.1016/j.it.2016.10.002
- Becker JC, Stang A, Schrama D, Ugurel S. Merkel cell carcinoma: integrating epidemiology, immunology, and therapeutic updates. Am J Clin Dermatol 2024; 25: 541–557. https://doi.org/10.1007/s40257-024-00858-z
- D‘Angelo SP, Lebbe C, Mortier L, Brohl AS, Fazio N, Grob JJ, et al. First-line avelumab in a cohort of 116 patients with metastatic Merkel cell carcinoma (JAVELIN Merkel 200): primary and biomarker analyses of a phase II study. J Immunother Cancer 2021; 9: e002646. https://doi.org/10.1136/jitc-2021-002646
- Nghiem P, Bhatia S, Lipson EJ, Sharfman WH, Kudchadkar RR, Brohl AS, et al. Durable tumor regression and overall survival in patients with advanced Merkel cell carcinoma receiving pembrolizumab as first-line therapy. J Clin Oncol 2019; 37: 693–702. https://doi.org/10.1200/JCO.18.01896
- Silk AW, Barker CA, Bhatia S, Bollin KB, Chandra S, Eroglu Z, et al. Society for Immunotherapy of Cancer (SITC) clinical practice guideline on immunotherapy for the treatment of nonmelanoma skin cancer. J Immunother Cancer 2022; 10: e004434. https://doi.org/10.1136/jitc-2021-004434
- Harjunpaa H, Guillerey C. TIGIT as an emerging immune checkpoint. Clin Exp Immunol 2020; 200: 108–119. https://doi.org/10.1111/cei.13407
- Banta KL, Xu X, Chitre AS, Au-Yeung A, Takahashi C, O‘Gorman WE, et al. Mechanistic convergence of the TIGIT and PD-1 inhibitory pathways necessitates co-blockade to optimize anti-tumor CD8(+) T cell responses. Immunity 2022; 55: 512–526 e519. https://doi.org/10.1016/j.immuni.2022.02.005
- Kawashima S, Inozume T, Kawazu M, Ueno T, Nagasaki J, Tanji E, et al. TIGIT/CD155 axis mediates resistance to immunotherapy in patients with melanoma with the inflamed tumor microenvironment. J Immunother Cancer 2021; 9: e003134. https://doi.org/10.1136/jitc-2021-003134
- Albrecht T, Brinkmann F, Albrecht M, Lonsdorf AS, Mehrabi A, Hoffmann K, et al. Programmed death ligand-1 (PD-L1) is an independent negative prognosticator in Western-World gallbladder cancer. Cancers (Basel) 2021; 13: 1682. https://doi.org/10.3390/cancers13071682
- Lonsdorf AS, Edelmann D, Albrecht T, Brobeil A, Labrenz J, Johanning M, et al. Differential immunoexpression of inhibitory immune checkpoint molecules and clinicopathological correlates in keratoacanthoma, primary cutaneous squamous cell carcinoma and metastases. Acta Derm Venereol 2024; 104: adv13381. Corrigendum: Acta Derm Venereol 2024; 104: 41934. https://doi.org/10.2340/actadv.v104.41934. https://doi.org/10.2340/actadv.v104.13381
- Hung AL, Maxwell R, Theodros D, Belcaid Z, Mathios D, Luksik AS, et al. TIGIT and PD-1 dual checkpoint blockade enhances antitumor immunity and survival in GBM. Oncoimmunology 2018; 7: e1466769. https://doi.org/10.1080/2162402X.2018.1466769
- Xiao Y, Chen P, Luo C, Xu Z, Li X, Liu L, et al. Discovery of a novel anti PD-L1 X TIGIT bispecific antibody for the treatment of solid tumors. Cancer Treat Res Commun 2021; 29: 100467. https://doi.org/10.1016/j.ctarc.2021.100467
- Thibaudin M, Limagne E, Hampe L, Ballot E, Truntzer C, Ghiringhelli F. Targeting PD-L1 and TIGIT could restore intratumoral CD8 T cell function in human colorectal cancer. Cancer Immunol Immunother 2022; 71: 2549–2563. https://doi.org/10.1007/s00262-022-03182-9
- Wu L, Mao L, Liu JF, Chen L, Yu GT, Yang LL, et al. Blockade of TIGIT/CD155 signaling reverses T-cell exhaustion and enhances antitumor capability in head and neck squamous cell carcinoma. Cancer Immunol Res 2019; 7: 1700–1713. https://doi.org/10.1158/2326-6066.CIR-18-0725
- Murakami D, Matsuda K, Iwamoto H, Mitani Y, Mizumoto Y, Nakamura Y, et al. Prognostic value of CD155/TIGIT expression in patients with colorectal cancer. PLoS One 2022; 17: e0265908. https://doi.org/10.1371/journal.pone.0265908
- Blessin NC, Simon R, Kluth M, Fischer K, Hube-Magg C, Li W, et al. Patterns of TIGIT expression in lymphatic tissue, inflammation, and cancer. Dis Markers 2019; 2019: 5160565. https://doi.org/10.1155/2019/5160565
- Rasmussen JH, Lelkaitis G, Hakansson K, Vogelius IR, Johannesen HH, Fischer BM, et al. Intratumor heterogeneity of PD-L1 expression in head and neck squamous cell carcinoma. Br J Cancer 2019; 120: 1003–1006. https://doi.org/10.1038/s41416-019-0449-y
- Schildhaus HU. [Predictive value of PD-L1 diagnostics]. Pathologe 2018; 39: 498–519. https://doi.org/10.1007/s00292-018-0507-x
- Schrama D, Peitsch WK, Zapatka M, Kneitz H, Houben R, Eib S, et al. Merkel cell polyomavirus status is not associated with clinical course of Merkel cell carcinoma. J Invest Dermatol 2011; 131: 1631–1638. https://doi.org/10.1038/jid.2011.115
- Hothorn T, Hornik K, van de Wiel MA, Zeileis A. Implementing a Class of Permutation Tests: The coin Package. Journal of Statistical Software 2008; 28: 1–23. https://doi.org/10.18637/jss.v028.i08
- Kim HS, Lee JY, Lim SH, Park K, Sun JM, Ko YH, et al. Association between PD-L1 and HPV status and the prognostic value of PD-L1 in oropharyngeal squamous cell carcinoma. Cancer Res Treat 2016; 48: 527–36. https://doi.org/10.4143/crt.2015.249
- Johnston RJ, Comps-Agrar L, Hackney J, Yu X, Huseni M, Yang Y, et al. The immunoreceptor TIGIT regulates antitumor and antiviral CD8(+) T cell effector function. Cancer Cell 2014; 26: 923-937. 26: 923–937. https://doi.org/10.1016/j.ccell.2014.10.018
- Chu X, Tian W, Wang Z, Zhang J, Zhou R. Co-inhibition of TIGIT and PD-1/PD-L1 in cancer immunotherapy: mechanisms and clinical trials. Mol Cancer 2023; 22: 93. https://doi.org/10.1186/s12943-023-01800-3
- Zhang P, Liu X, Gu Z, Jiang Z, Zhao S, Song Y, et al. Targeting TIGIT for cancer immunotherapy: recent advances and future directions. Biomark Res 2024; 12: 7. https://doi.org/10.1186/s40364-023-00543-z
- Ishihara S, Iwasaki T, Kohashi K, Kawaguchi K, Toda Y, Fujiwara T, et al. Clinical significance of signal regulatory protein alpha and T cell immunoreceptor with immunoglobulin and immunoreceptor tyrosine-based inhibition motif domain expression in undifferentiated pleomorphic sarcoma. J Cancer Res Clin Oncol 2023; 149: 2425–2436. https://doi.org/10.1007/s00432-022-04078-y
- Becker JC, Beer AJ, DeTemple VK, Eigentler T, Flaig M, Gambichler T, et al. S2k Guideline - Merkel cell carcinoma (MCC, neuroendocrine carcinoma of the skin) - Update 2022. J Dtsch Dermatol Ges 2023; 21: 305–320. https://doi.org/10.1111/ddg.14930
- Hansen UK, Church CD, Carnaz Simoes AM, Frej MS, Bentzen AK, Tvingsholm SA, et al. T antigen-specific CD8+ T cells associate with PD-1 blockade response in virus-positive Merkel cell carcinoma. J Clin Invest 2024; 134: e177082. https://doi.org/10.1172/JCI177082